






 12,5 мкм. Функция микроворсинок пока неизвестна.
12,5 мкм. Функция микроворсинок пока неизвестна.
Внутренние членики (сегменты) палочек имеют цилиндрическую форму. Гистологически различают две части внутреннего членика: эозинофильную наружную, называемую эллипсоидной частью, и внутреннюю базофильную, называемую миоидной частью (рис. 3.6.16— 3.6.19). Тинкториальные свойства этих двух областей изменяются в зависимости от метаболической активности фоторецептора. Эллипсоидная часть окрашивается эозинофильно в связи с наличием в ней большого количества митохондрий. В одной палочке можно найти до 600 митохондрий. Цитоплазма также содержит гладкий эндоплазматический ретикулум, нейро-трубочки, свободные рибосомы и гранулы гликогена. Базофилия миоидной части зависит от большой концентрации в ней свободных рибосом. Миоидная область является центром белкового синтеза. По этой причине в ней располагаются шероховатая эндоплазматическая сеть, аппарат Гольджи, микротрубочки, микрофила-менты и гранулы гликогена. Таким образом, основной функцией этой части фоторецептора является обеспечение метаболических и синтетических функций клетки.
Наружные сегменты (членики) колбочек имеют различное строение в различных участках сетчатки. В области зубчатой линии и по периферии сетчатки они короткие и конические, а в fovea centralis продолговатые, напоминают наружные сегменты палочек (рис. 3.6.16—3.6.18).
Ультраструктурными исследованиями установлено, что наружный сегмент колбочки имеет больше дисков (1000—1200), чем наружный сегмент палочки. Междисковые пространства палочки более широкие (в колбочках — 3,5 мкм, в палочках— 16,5 мкм).
В отличие от дисков палочек диски колбочек соединены между собой и прикрепляются к плазматической мембране.
Внутренние сегменты (членики) колбочек. Наружные и внутренние членики колбочек связаны друг с другом посредством тонкого цитоплазматического перешейка, содержащего видоизмененную ресничку. Они изменяются в зависимости от их топографического расположения. В центральной ямке (fovea centralis) они более длинные и узкие. Ультраструктурная организация внутреннего сегмента палочек и колбочек одинаковая, за исключением того, что в колбочках значительно больше митохондрий (200—300 на срезе).
Наружная поверхность миоидной части палочек и колбочек покрыта волосоподобными цитоплазматическими отростками мюллеровс-ких клеток, формирующих «корзины Шульца». Благодаря этому никакого контакта между смежными клетками нет [1008]. Отростки мюл-леровских клеток участвуют также в регуляции состава внеклеточной среды фоторецепто-
ров и служат для жесткой пространственной фиксации палочек и колбочек.
Наружная пограничная мембрана. При световой микроскопии видно, что наружная пограничная мембрана (рис. 3.6.1) отделяет слой палочек и колбочек от подлежащего наружного ядерного слоя сетчатки. Она простирается от диска зрительного нерва до зубчатой линии, где превращается в базальную пластинку, расположенную между пигментированными и беспигментными частями ресничного эпителия. Наружная пограничная мембрана представляет собой не что иное, как скопление в одной плоскости терминальных пластинок (zonulae adhe-rentes), расположенных между мюллеровскими клетками и фоторецепторами, между смежными мюллеровскими клетками и, редко, между соседними фоторецепторами.
Наружная пограничная мембрана, таким образом, не является истинной мембраной. Через нее проходят небольшие молекулы. Главной функцией мембраны является обеспечение функционирования избирательного барьера на пути питательных веществ, которые проходят между рядом расположенными мюллеровскими клетками, а также стабилизация положения фоторецепторов.
Наружный ядерный слой. Наружный ядерный слой находится кнутри от наружной пограничной мембраны и содержит тела и ядра фото-рецепторных клеток (рис. 3.6.1). В зависимости от участка сетчатки ширина этого слоя изменяется, прежде всего, из-за изменения числа рядов ядер.
С назальной стороны диска наружный ядерный слой имеет толщину 45 мкм и состоит из 8—9 рядов ядер. С височной стороны он состоит только из четырех рядов ядер, истончаясь до 22 мкм. В желтом пятне наличие 10 рядов ядер колбочек увеличивают ширину наружного ядерного слоя до 50 мкм. В области зубчатой линии наружный ядерный слой состоит только из одного слоя ядер колбочковых клеток, которые плотно прилежат к наружной пограничной мембране и четырем рядам ядер палочек, расположенным кнутри от них. Толщина ядерного слоя при этом приблизительно 27 мкм.
Ядра колбочек овальные и имеют диаметр 5—7 мкм. Расположены они на 3—4 мкм кнутри от наружной пограничной мембраны. Ядра палочек также овальные, диаметром 5,5 мкм.
Цитоплазма обоих типов клеток скудная. Тела палочек и колбочек окрашиваются по-разному. При использовании метода Унна тело палочек не окрашивается, а колбочки окрашиваются в интенсивно синий цвет. Используя трехцветный метод Маллори, после фиксации сетчатки жидкостью Ценкера можно четко дифференцировать центральную ямку. Центральная ямка окрашена в интенсивно красный цвет. Это связано с тем, что методом Маллори окрашиваются только колбочки.
Сетчатка
 Наружный сетчатый (плексиформный) слой (рис. 3.6.1) является местом соединения первого и второго нейронов, т. е. местом передачи информации от первого нейрона (фоторецептора) второму (биполярной клетке). Помимо указанных клеток в нем располагаются ассоциативные нейроны (горизонтальная клетка).
Наружный сетчатый (плексиформный) слой (рис. 3.6.1) является местом соединения первого и второго нейронов, т. е. местом передачи информации от первого нейрона (фоторецептора) второму (биполярной клетке). Помимо указанных клеток в нем располагаются ассоциативные нейроны (горизонтальная клетка).
Две трети слоя состоит из внутренних волокон фоторецепторов, окруженных отростками мюллеровских клеток. Треть слоя состоит из дендритов биполярных и горизонтальных клеток, а также отростков мюллеровских клеток. Наружный плексиформный слой наиболее толстый в области желтого пятна (51 нм). Состоит он из косо идущих волокон, отклоняющихся от желтого пятна. Этот слой также известен как слой волокон Хенле.
Внутренние волокна в наружном плексифор-мном слое представляют собой аксоны палочек и колбочек. Диаметр аксона палочки приблизительно в четыре раза больше, чем у колбочки. Они содержат типичные органоиды — единичные митохондрии, немного свободных рибосом, гладкий эндоплазматический ретикулум, гранулы гликогена и плотно упакованные микротрубочки.
Синаптическая связь палочек со вторым нейроном происходит при помощи овальных расширений цитоплазмы диаметром 1 мкм. Называются они сферулами.
Синапсы колбочек отличаются. Эти отличия сводятся к тому, что колбочки образуют так называемую «ножку», т. е. ножкоподобное утолщение окончания цитоплазматического отростка колбочки. «Ножка» больше, чем сфе-рула (7—8 мкм в парафовеолярной области и 5 мкм в области фовеа). Теперь мы более подробно остановимся на синаптических связях этого слоя.
Синапсы палочек. Синаптический комплекс палочек состоит из указанной выше пресинап-тической сферулы, синаптической ленты и постсинаптических отростков, принадлежащих горизонтальным или биполярным клеткам (рис. 3.6.20—3.6.23). Сферулы содержат многочисленные пресинаптические пузырьки, а также митохондрии и нейротрубочки. Плотность пре- и постсинаптической мембраны увеличивается вблизи синаптической щели (ширина синаптической щели 15 мкм). Перпендикуляр, проходящий через пресинаптическую мембрану, называется синаптической лентой, состоящей из трех электронноплотных слоев, каждый из которых имеет толщину 12 мкм. Отделяется она светлой зоной, имеющей толщину 40 мкм, и окружена ореолом пузырьков. Сферулы палочек содержат только две синаптические ленты, которые ассоциируются с двумя боковыми элементами, являющимися терминалами аксонов горизонтальных клеток, и двумя дендри-тами биполярных клеток палочек [263, 593] (рис. 3.6.22).
С одной сферулой палочки может входить в контакт несколько различных горизонтальных клеток (1—4 клетки). Различают два основных типа контактов — с телодендритами горизонтальной клетки и дендритом биполярной клетки. Каждая сферула входит в контакт с 4 биполярными клетками. В то же время каждая биполярная клетка контактирует с 50 палочками (вне фовеолы) и с несколькими сотнями палочек по периферии сетчатки [596].
Эти различия в характере межнейронных связей соответствуют различиям в разрешающей способности зрительной системы.
Синапсы колбочек. «Ножка» колбочки пирамидальной формы. Синаптические вдавления на «ножке» объединяют одновременно три нейрона, контактирующие в то же самое время и между собой. Подобная структура получила название «триада» [263, 593, 1008] (рис. 3.6.20, 3.6.21). Центральный аксон триады принадле-
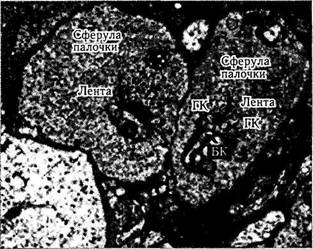

Рис. 3.6.20. Ультраструктурные особенности сферул палочек (а) и «ножек» колбочек (б) (по Kolb, 1998):
НПС — наружный плексиформный слой; ГК—горизонтальная клетка; БК—биполярная клетка палочки; ИБК—инвагинирую-щая биполярная клетка; ПБК — плоская биполярная клетка
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 | |

|
| Колбочки («ножка») |
Палочки («сферулы»)
Рис. 3.6.21. Схематическое изображение синаптичес-ких тел палочки и колбочки:
/ — палочковые биполярные клетки; 2 — карликовая биполярная клетка; 3 — плоская биполярная клетка; 4 — горизонтальная клетка. Отмечается наличие контакта между сферулой палочки и «ножкой» колбочки. Синаптические тела палочки и колбочки соединяются непосредственно, а также при помощи горизонтальной клетки
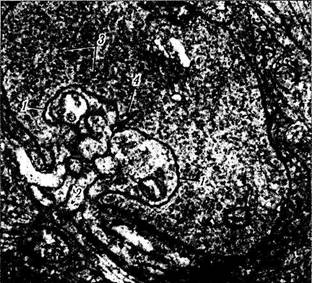
Рис. 3.6.22. Электроннограмма сферулы палочки:
/ — латеральные отростки горизонтальной клетки; 2 — отростки
биполярной клетки; 3 — синаптические пузырьки; 4 — синапти-
ческая лента
жит биполярной клетке. Это аксон может входить в контакт с той же самой колбочкой в 10—25 различных точках [172, 186]. Два дендрита с обеих сторон триады исходят из различных горизонтальных клеток. Хотя только одна биполярная клетка входит в контакт с одной «ножкой» колбочки, контакт существует со многими горизонтальными клетками, число которых обычно б—8. Такая «ножка» имеет также множество маленьких поверхностных вдавлений (так называемых базальных соеди-

Рис. 3.6.23. Особенности синаптических связей палочек и колбочек с биполярными клетками и биполярных клеток с ганглиозными:
видно, что одна биполярная клетка получает информацию от нескольких палочковых фоторецепторов и только от одной колбочки
нений), контактирующих с плоской диффузной биполярной клеткой [453, 505, 637]. Подобный тип синапсов биполярных клеток формируется сразу с шестью колбочками [661]. Базальные соединения представляют собой классические эксцитаторные (возбуждающие) синапсы и функционируют подобно щелевым контактам.
Присутствие многочисленных десмосом между отростками клеток наружного плекси-формного слоя (десмосомы) препятствует свободному распространению в сетчатке метаболитов, жидкостей и экссудата.
Помимо биполярных и горизонтальных клеток, фоторецепторы контактируют и между собой. Палочки контактируют с палочками и колбочками. Происходит это благодаря так называемым щелевым контактам. От «ножки» колбочки отходят тонкие отростки, которые подходят к сферулам палочек и «ножкам» других колбочек. В тех местах, где эти отростки (называемые телодендритами) образуют щелевой контакт, формируется «электрический контакт», т. е. происходит передача информации без использования нейротрансмиттера [742, 774, 870]. 3—5 подобных контактов определяется на одной сферуле палочки, образованной телодендритами колбочки. Одна «ножка» колбочки может иметь до 10 контактов с соседними палочками. «Ножки» S-колбочек («синих») не содержат такого большого количества контактов [43]. По этой причине S-колбочки довольно изолированы.
Сетчатка


|
Функциональное значение прямой электрической связи между различными типами фоторецепторов не совсем понятно. Первоначально многие исследователи предполагали, что такие связи разрушают пространственную интеграцию фоторецепторов и соответственно возможность анализа функционирования цветового зрения, «смешивая» информацию, получаемую от палочек и колбочек. Тем не менее на основании многочисленных физиологических экспериментов установлено, что колбочки благодаря этим связям могут нести информацию палочек. Это, при определенных условиях, может иметь большое физиологическое значение [742, 773, 975, 999]. При этом изучены интимные механизмы этого процесса, правда, с использованием экспериментальных животных.
Внутренний ядерный слой. Внутренний ядерный слой состоит из 8—12 рядов плотно упакованных ядер биполярных, горизонтальных, амакриновых, межплексиформных и мюл-леровских клеток. При световой микроскопии можно различить четыре слоя, преимущественно содержащих тот или иной клеточный тип:
1. Слой горизонтальных клеток (наиболее
наружный).
2. Слой биполярных клеток (наружный про
межуточный слой).
3. Слой мюллеровских клеток (внутренний
промежуточный).
4. Слой амакриновых и межплексиформных
клеток (самый внутренний).
Горизонтальные клетки (рис. 3.6.24— 3.6.25; 3.6.26, см. цв. вкл.). Отростки горизон-

Рис. 3.6.24. Особенности строения тел и дендритного поля различных типов горизонтальных клеток человека. Световая микроскопия (импрегнация серебром) (по Kolb, 1998)


Рис. 3.6.25. Схематическое изображение различных типов горизонтальных клеток:
а — горизонтальная клетка, контактирующая с колбочковым фоторецептором; б —горизонтальная клетка, контактирующая с палочковым фоторецептором; s — схематическое изображение характера контакта горизонтальных клеток различного типа в плоскости сетчатки
тальных клеток, в отличие от биполярных, образуют сеть, расположенную в горизонтальной плоскости и объединяющую фоторецепторы различных участков сетчатки.
Наибольшее количество горизонтальных клеток в области центральной ямки. Постепенно по мере продвижения к периферии сетчатки их число снижается. Горизонтальные клетки имеют короткие отростки, а аксон не ветвится вблизи тела клетки (на протяжении 200— 300 мкм). Длина аксона может достигать 2 мм.
В зависимости от размера клетки, особенностей строения синапсов между дендритами и аксонами, а также площади дендритного поля различают три типа горизонтальных клеток. Обозначаются они как клетки типов HI, НИ

