




Структурною і функціональною одиницею спадковості, згідно з сучасними уявленнями, є ген. Місце локалізації генів – хромосоми. У всіх багатоклітинних організмів нараховуються тисячі генів, тоді як число хромосом досить обмежен (звичайно від декількох до небагатьох десятків). Отже кожна хромосома повинна містити багато генів, що і було доведено численними генетичними і цитологічними дослідженнями.
Відомо, що хромосоми успадковуються як єдине ціле. Тому і гени, локалізовані в одній хромосомі, як правило, успадковуються разом. Гени, локалізовані в одній хромосомі, складають одну групу зчеплення. А спільне успадкування генів, які складають групу зчеплення, називається зчепленим успадкуванням генів. Явище зчеплення ознак (генів) відкрили в 1906 р. В.Бетсон і Р.Пенкет, пізніше цим питанням займався цитолог У.Сеттон. Проте розкриття суті цього явища стало можливим завдяки працям Т.Моргана та його співробітників (А.Стертевант, Г.Меллер, К.Бреджес). Закономірності зчепленого успадкування викладені у створеній цими вченими хромосомній теорії спадковості.
Основні положення хромосомної теорії спадковості та їх тлумачення, необхідні для вирішення задач:
Гени локалізуються в хромосомах. Кожний ген у хромосомі займає певну ділянку — локус.
Гени в одній хромосомі згідно з хромосомною теорією розташовані лінійно (по довжині хромосоми).
Гени, локалізовані в одній хромосомі, звичайно, успадковуються разом і складають одну групу зчеплення.
Алельні гени локалізовані в певних ділянках гомологічних хромосом, тому групу зчеплення складають дві гомологічні хромосоми. Отже, на основі принципу гомологічності хромосом, число груп зчеплення звичайно відповідає кількості пар хромосом (гаплоїдній кількості хромосом каріотипу). Наприклад, у мушки дрозофіли диплоїдний набір – 8 хромосом і тому 4 групи зчеплення, у людини 46 хромосом і тому 23 групи зчеплення і т.д. Таке співвідношення кількості груп зчеплення і гаплоїдного набору хромосом повністю простежується у гомогаметної статі і дещо не збігається у гетерогаметної статі цього ж виду організмів.
При розв’язанні задач щодо визначення кількості груп зчеплення звичайно приймається до уваги диплоїдний набір хромосом, характерний для того чи іншого виду без урахування статі.
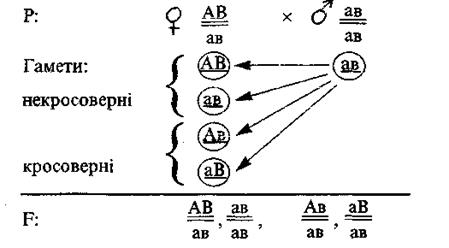
З метою полегшення розв’язування задач використовують особливий запис генотипу і гамет, який дає змогу в письмовій формі диференціювати зчеплені ознаки від ознак, гени яких знаходяться в різних хромосомах. Наприклад, генотип дигетерозиготи АаВв можна записати як  або
або  .
.
Дві паралельні лінії символізують гомологічні хромосоми, а комбінації генів над і під цими лініями – групи зчеплення, успадковані організмом від матері (верхня) і від батька (під низом). Гамети з вихідними групами зчеплення (незмінною комбінацією генів) називають некросоверними, а гамети з новими комбінаціями генів – кросоверними. У запису гамет теж використовують символічні хромосоми. Наприклад, в особини з генотипом за умов повного зчеплення утворюються лише некросоверні гамети (АВ) і (ав), а за умов неповного зчеплення, окрім уже записаних сортів гамет, утворюються ще і кросоверні гамет (Ав) і (аВ). Отже, при повному зчепленні дигетерозиготний організм продукує два сорти гамет, а при неповному зчепленні – чотири сорти гамет.
Такий вид запису генотипів і гамет використовується для позначення зчеплених ознак, гени яких локалізовані в аутосомах.
При зчепленому успадкуванні можливе виникнення нових комбінацій генів у гаметах, що пояснюється явищем кросинговеру. Кросинговер (перехрест хромосом) відбувається у профазі першого поділу мейозу, під час якого гомологічні хромосоми (точніше, прилеглі під час кон’югації хроматиди) обмінюються своїми ділянками і утворюються рекомбінантні (кросоверні) хромосоми. Рекомбінантні гомологічні хромосоми містять ділянки (гени) як батьківських, так і материнських хромосом. Наприклад, дигетерозиготний організм з генотипом (за умов неповного зчеплення) може продукувати чотири типи гамет: (АВ), (ав), (Ав), (аВ). Гамети типу (Ав) і (аВ) утворилися внаслідок кросинговеру, тобто між хроматадами гомологічних хромосом відбувся обмін певними ділянками, що призвело до рекомбінації генетичного матеріалу і появи відмінного від вихідної форми сполучення генів. Схематично це можна зобразити так:

Кожна із утворених хроматид (надалі – самостійні хромосоми) потрапляє в окрему гамету і утворюється чотири їх типи. Проте, співвідношення цих гамет на відміну від вільного комбінування буде нерівним. Пояснюється це тим, що кросинговер відбувається не з постійною частотою і тому рекомбінантні гамети завжди складають не менше 50%.
Частота кросинговеру прямо пропорційна відстані між генами в групі зчеплення: чим далі розташовані один від одного гени, тим частіше спостерігається кросинговер. Отже, чим більша відстань між генами в групі зчеплення, тим вища ймовірність перехресту і розвитку кросоверних гамет і навпаки. Тому некросоверних гамет завжди більша кількість, ніж кросоверних.
Особини, що розвиваються з кросоверних (рекомбінантних) гамет, називають кросоверами (рекомбінантами). Виникають рекомбінантні особини з такою самою ймовірністю, як і кросоверні гамети. І, відповідно, особини, що розвиваються з некросоверних гамет, називають некросоверами (нерекомбінантами).
Процент кросинговеру є мірою відстані між генами. Для визначення цієї відстані використовують винятково аналізуюче схрещування, тобто схрещування з гомозиготним за рецесивними ознаками організмом. При такому схрещуванні процент рекомбінантів дорінює як частоті кросоверних гамет, так і відстані між відповідними генами. Наприклад, від аналізуючого схрещування дигетерозиготи одержано 12% кросоверів.
Схема схрещування:
| некросоверні (нерекомбінанти) Всього 100% - 12% = 88%, по 44% кожного типу гамет або класу генотипів і фенотипів | кросоверні (рекомбіновані) Всього: 12 %, по 6 % кожного типу гамет або класу генотипів і фенотипів |
За одиницю відстані між генами у хромосомі прийнята 1 морганіда, яка дорівнює 1% кросинговера. Отже, 12 % кросоверів означають, що дигетерозигота утворила 12% кросоверних гамет і що відстань між генами А і В (а також а і в, А і в, а і В) становить 12% або і 2 морганід.
Для розрахунку відносної відстані між генами, які локалізовані в одній хромосомі, використовують формулу:
 де
де
X – процент кросоверів від аналізуючого схрещування;
(а+с) – кількість кросоверів;
n – загальна кількість потомства.
За частотою кросинговеру можна визначити відстань між зчепленими генами, що дозволяє складати генетичні карти. Генетичні карти хромосом – це схеми, на яких вказано порядок розташування генів у хромосомі і відносна відстань між ними. Для складання генетичної карти хромосом будь-якого організму досліджують успадкування не менше трьох генів групи зчеплення.
Розв’язок задач з даного розділу базується на викладеному вище матеріалі. Слід пам’ятати, що у самця дрозофіли, самки шовковичного шовкопряда кросинговер не спостерігається.

