




Проведенные исследования и опыты на бактериях и вирусах, растениях и животных убедительно доказали о связи наследственности организма с хромосомами.
Хромосомы несут в себе информацию о свойствах и признаках организма. Эта наследственная информация через хромосомы и клетки передается от одного поколения к другому. Таким образом, клетка является генетической системой, где сосредоточена вся наследственная информация. Ядро является хранилищем наследственной информации, а местом реализации наследственной информации – является цитоплазма.
По химическому составу хромосомы состоят из нуклеиновых кислот (ДНК, РНК и белка типа гистонов).
Нуклеиновые кислоты открыты в 1868г. Ф. Мишером, который выделил их из клеточных ядер человека, а затем из спермиев лососей.
Найденное вещество назвали нуклеином, подчеркнув их принадлежность к клеточному ядру. ДНК и РНК обладает способностью воспроизводить себе подобные и контролировать синтез строго специфических белков или ферментов.
ДНК и РНК принадлежит ведущая роль в передаче наследственной информации. Они имеют различные функции, различное химическое строение и места расположения.
ДНК – ее основная масса находится в хромосомах ядра. Сравнительно небольшая часть ДНК, около 5;% локализованы в митохондриях.
РНК – рибонуклеиновая кислота, ее основная масса находится в цитоплазме.
ДНК – гигантская молекула, она компактно упакована в хромосоме. В каждой хромосоме содержится всего одна молекула ДНК.
ДНК – по своей природе представляет собой сложный биологический полимер, имеет организованную систему, высокий молекулярный вес от 5 до 40млн. ед.может доходить до 100 млн. ед. Линейные размеры хромосомы примерно в 10000 раз меньше, чем содержащейся в ней ДНК. Модель структуры ДНК была предложена в 1953году Дж. Уотсоном и Ф. Криком.
В бактериальной клетке длина молекул ДНК достигает 1см, а в клетке человеческого тела более 1м (длина ДНК в 46 хромосомах).
У взрослого человека примерно 5х 1013 клеток, так что общая длина молекул ДНК в организме 1011 км (почти в тысячу раз больше расстояние от Земли до Солнца).
В состав ДНК входит сахар-дезоксирибоза фосфорные остатки и азотистые основания.
Азотистые основания 2х типов:
 аденин (А)
аденин (А)
 пуриновые
пуриновые
гуанин (Г)
 тимин (Т)
тимин (Т)
 пиримидиновые
пиримидиновые
цитозин (Ц)
Комплекс: фосфорной кислоты
сахара дезоксирибозы (углевод)
азотистые основания
 Ф-Д образуют нуклеотид
Ф-Д образуют нуклеотид
׀
А
Комплекс: Д сахара дезоксирибозы
׀
А азотистые основания образуют нуклеозид
Название нуклеотида обычно происходит от входящего в его состав нуклеозида, порядкового номера атома пентозы, к которому присоединен остаток фосфорной кислоты, или называют, как кислоту, по азотистому основанию. Название нуклеозида определяется содержащимся в нем азотистым основанием. Так, рибонуклеозид, имеющий в составе молекулы аденин, называют аденозином, гуанин – гуанозином, Тимин- тимидином, цитозин – цитидином, урацил – уридином. Дезоксирибонуклеозид: дезоксиаденозин, дезоксигуанозин, дезоксицитидин, тимидин.
Химический анализ показал, что в ДНК любых организмов количество А (аденина) всегда точно соответствует количеству Т (тимина), а количество гуанина – количеству Ц (цитозина) А=Т, Ц=Г. Таким образом, сумма пуриновых оснований (А+Г) равна сумме пиримидиновых оснований (Т+Ц). Эта зависимость была впервые установлена американским биохимиком Э. Чаргаффом А+Г / Т+Ц = 1 (правило нуклеиновых отношений)
В ДНК азотистые основания соединены друг с другом водородными связями в строго определенном сочетании, аденин всегда только с тимином, цитозин только с гуанином. Связь, при которой друг с другом соединяются лишь строго определенные химические вещества А↔Т и Г↔Ц, называется комплементарной.
Для каждого вида и индивидуума имеется специфическая молекула ДНК или наследственность.
Биологическое значение ДНК – в том, что на ней зашифрована вся наследственная информация любого организма.
Индивидуальность наследственной информации обеспечивается числом и порядком чередований азотистых оснований.
Каждый индивид имеет свою ДНК (вот в этом состоит барьер несовместимости - при пересадке кожи или других каких органов от одного человека другому, ДНК донора не совпадает с ДНК реципиента (больного), происходит отторжение даже через несколько лет.
В соматических клетках и ДНК в два раза больше, чем в половых, потому, что в соматический диплоидный набор хромосом, а в половых – гаплоидный.
ДНК всех живых существ в целом устроена одинаково, однако у разных видов установлено разное отношение молекулярной суммы А+Т к молекулярной сумме Г+Ц. Это отношение называют коэффициентом видовой специфичности введенной Д. Уотсоном. Видовая специфичность выражается долей в ней пар ГЦ. Эта доля неодинакова у разных видов.
А+Т
Г+Ц = 0,75 – данное соотношение у крупного рогатого скота;
0,72 – у кур;
1,04 – кишечной палочки;
0,66 – у человека.
Рентгеноструктурный анализ показал, что молекула ДНК состоит из двух цепей, эти спирально закрученные цепи (вокруг собственной оси) могут вытягиваться и складываться.
Наружный остов этих нитей состоит из чередующихся молекул сахара и фосфора, а внутреннюю часть составляют пары азотистых оснований, соединенных между собой водородными связями между аденином (А) и тимином (Т) двойная водородная связь. Цитозином (Ц) и гуанином (Г), тройная (А=Т) (Г=Ц). Молекула ДНК способна к автосинтезу (самоудвоению) – репликации.
Синтез ДНК. Этот процесс проходит в период клеточного покоя. ДНК находится в хромосомах и репликация её происходит перед каждым удвоением хромосом и делением клетки.
Репликация начинается с одного конца молекулы ДНК (в определенной точке старта) в хромосомах эукариот несколько точек. Спиралевидная ДНК раскручивается (расплетается вдоль оси, под действием комплекса ферментов) – главное из которых получил название ДНК – полимераза. Водородные связи разрываются на отдельных участках образуют вилку репликации.
Комплементарные нити разъединяются и каждая нить становится матрицей на которой проходит синтез и образование дочерних нитей.
У эукариот на каждой нити ДНК процесс репликации идет неодинаково. Репликация на материнской цепи, идущей от точки старта, идет ввиде сплошной линии, получившей название лидирующей.
Лидирующая нить – синтезируется при участии фермента ДНК – полимераза. Синтез «запаздывающей» нити идет сложней с участием комплекса ферментов, в противоположном направлении синтеза лидирующей нити. Вначале образуются отрезки-реплики новой дочерней нити ДНК. Они называются по имени открывшего их японского ученого Р. Оказаки. После завершения синтеза фрагменты Оказаки соединяются при помощи фермента лигазы в общую полинуклеотидную цепочку.
Чтобы доказать, что на молекуле ДНК записана вся наследственная информация проводился опыт на вирусах и бактериях и с помощью явлений трансформации, трансдукции и коньюгации доказано, что вся наследственная информация записана на ДНК.
Строение и типы РНК
РНК – рибонуклеиновая кислота состоит из фосфорного остатка, сахара – рибозы и азотистого основания – А, Г, Ц,,У. Вместо тимина Т-У (урацил).
Молекула РНК состоит из одной цепи и имеет различный молекулярный вес. Синтез РНК происходит на молекуле ДНК. Таким образом, молекула ДНК является «матрицей» для синтеза РНК. В синтезе участвует и специальный фермент РНК – полимераза.
Молекула ДНК расщепляется на две нити, но не по всей длине, а на определенном участке.
При синтезе образуются гибридные двухцепочные структуры ДНК/ РНК и существует до тех пор, пока не достигнет определенного соотношения. После чего РНК сходит с молекулы ДНК и попадает в цитоплазму через поры ядра.
Биологическое значение РНК – является переносчиком информации с ДНК к месту синтеза белка, на рибосомы.
Существует 3 вида РНК: 1вид – информационная (матричная) и-РНК или м-РНК, они имеют сотни или тысячи нуклеотидов.
Размеры молекул РНК различны, но в общем они меньше молекул ДНК
РНК считывает и переносит наследственную информацию из ядра с участка ДНК в цитоплазму (рибосомы). Ее количество не велико 5% от всего РНК, что содержится в клетке.
II вид тРНК- транспортная. Она представлена несколькими десятками форм. Функция – т-РНК – доставлять аминокислоты к месту синтеза белка – рибосомам, где каждая аминокислота имеет свою транспортную РНК.
Нить т-РНК имеет специфическую форму, напоминающую трехлопастный лист клевера.
Одна молекула т-РНК содержит в среднем 76-85 нуклеотидов. В клетке т-РНК – 10-15% от всех РНК.
III вид р-РНК – рибосомальная. Она входит в состав рибосом и служит как бы каркасом рибосом, определяющего их морфологию. Рибосомная РНК накапливается в ядрышках клетки и через поры ядра поступает в цитоплазму. Она составляет 80% от всей массы. Если и-РНК нестабильна, быстро обновляется, то р-РНК стабильна и раз образовавшиеся остается более или менее постоянным элементом клетки.
Таким образом, нуклеиновым кислотам ДНК и РНК принадлежит ведущая роль в передаче наследственной информации.
Строение и действие гена
Впервые доказал единицу наследственности Г. Мендель, он назвал ее наследственным фактором. Позже Иогансен предложил назвать генами.
С открытием хромосомной теории наследственности Морган установил, что гены в хромосоме расположены в линейном порядке и гены рассматривали как неделимые единицы наследственности.
Большой вклад в теорию строения гена внесли А.С. Серебровский и Н.П. Дубинин, 1920г. на основании своих исследований они создали центровую теорию строения гена. Согласно которой ген имеет определенную протяженность в хромосоме и состоит из отдельных единиц – центров. Эти центры способны мутировать, т.е. изменяться. Мутации могут захватывать не обязательно полностью весь ген, а проходить в отдельных его участках или захватывать несколько центров одновременно. Такой сложный ген получил название базигена.
Дробимость гена была доказана ученым. Наиболее сильное влияние на современное представление на современное представление о гене оказал американский физик и генетик С. Бензер. Он доказал, что ген состоит из 3-х линейно расположенных субъединиц, которые он назвал цистрон, мутон и рекон.
Цистроном – был назван участок ДНК, кодирующий первичную структуру одной полипептидной цепи или одной молекулы ДНК. Цистрон занимает в ДНК довольно длинные отрезки.
Кроме цистронов, ген включает участки способные мутировать. Они называются сайтами (мутон).
Мутон (сайт) – наименьший участок молекулы ДНК, или участок гена способный изменяться (мутировать). Раньше предполагали, что в состав мутона входит 5 нуклеотидов, а в настоящее время утверждают – 1 пару нуклеотидов, а в некоторых случаях, возможно и больше.
Рекон – участок гена, который не делится кроссинговером и он существует как единое целое. Предполагают, что в состав рекона входит несколько нуклеотидов.
Современное понятие гена – участок молекулы ДНК отвечающий за синтез определенного белка или фермента или самовоспроизводящий участок ДНК, контролирующий последовательность аминокислот в одной из полипептидной цепи белковой молекулы.(Ген-это основная физическая и функциональная единица наследственности, несущая информацию от одного поколения другому)
Ген является дискретной единицей наследственной информации, это локус (участок) хромосомы, оказывающий специфическое влияние на развитие организма. Одна молекула ДНК – это тысячи различных генов, но последовательность нуклеотидов каждого гена не повторяется ни в одном другом гене.
В состав гена входят низшего порядка – нуклеотиды. Любой ген имеет определенную величину, выраженную числом нуклеотидов и молекулярной массой.
Величина гена – связана с размером того белка, который образуется под его контролем. В среднем считают, что в белок входит 300-500 аминокислот. Если учесть, что молекулярная масса одной пары нуклеотидов равна 660 ед., а ген среднего размера состоит из 1500 нуклеотидных пар, то молекулярная масса гена выразится величиной 660 х 1500 более 1 млн. Ген занимает примерно одну десятитысячную часть хромосомы. Как элемент наследственности ген входит в непрерывную линейную структуру хромосом.
Каждый ген действует в системе целостного генотипа на ряд признаков. Ген является биологической единицей, он взаимодействует с цитоплазмой и изменяет ее, в тоже время цитоплазма действует на ген.
Свойства гена:
1.Дискретность, целостность т.к. он состоит из набора нуклеотидов – он представляет единое целое.
2.Аллельность – хорошо видна при наследовании тех или иных признаков, он может быть доминантным и рецессивным.
3.Постоянство гена подтверждается стабильностью нуклеиновых кислот.
Например: у некоторых простейших имеется кремневый скелет, который сохраняется в поколениях без существенных изменений сотни миллионов лет. Мутации генов, происходящие в отрезке времени не нарушают закона постоянства.
Из общих свойств гена следуют его частные свойства:
аллелизм – один ген может изменяться несколько раз (окраска меха у кроликов, глаза у мухи дрозофилы).
Кроме свойств гены обладают действием
1.Аддитивное
2.Плейотропное
3.Полимерное
4.Модифицирующее
5.Летальное
6.Мутаторное
Аддитивные – гены оказывают суммарное действие на признак.
Плейотропное – действие одного гена не на один, а на несколько признаков.
Пример: 1.Желтая мышь, у которой есть ген, вызывающей желтую окраску. У таких мышей, помимо желтой окраски обнаруживается склонность к ожирению, утяжелению костяка и определенная устойчивость к заболеванию раком молочной железы.
2.Ген F вызывающий у кур курчавость их оперения, обуславливает также нарушение деятельности гормональной, пищеварительной, сердечно-сосудистой систем организма и недоразвитие яичника, что является причиной низкой яйценоскости кур.
Для селекционера плейотропное действие гена нежелательное явление оно затрудняет отбор практически полезных форм. Так положительное явление одного признака может одновременно сопровождаться отрицательным изменением другого.
Полимерное – на один признак действует много генов. Все хозяйственно- полезные признаки являются полимерными. Такие как молочность скота, яйценоскость кур, масса и рост животного, скороспелость животного и др.
Модифицирующие - гены, не проявляющие собственного, но оказывающие влияние на эффект действия других генов. Гены модификаторы не изменяются, а ослабляют или усиливают действие других генов (пегость у лошадей, черно-пестрая масть крупного рогатого скота).
Летальные гены – ген, вызывающий в гомозиготном состоянии гибель организма. При разных летальных генах организмы гибнут на различных стадиях развития, иногда наблюдаются аборты, рождение уродов, однако чаще они гибнут на ранних стадиях.
Обычно летальное действие таких генов рецессивно, т.е. когда они находятся в гомозиготном состоянии. При возникновении мутаций с доминирующем летальным действием организм погибает не давая потомства, следовательно, наблюдать не удается.
Пример летального действия:
У каракульских овец шкурки ценятся с серебристо-серой окраской
♂Аа х ♀Аа (ширази)
сер. сер.
АААа Аа аа
сер. сер. черн.
↓
25 % ягнят гибнут при переходе на грубый корм (заболевали тимпанитом и погибали).
Мутаторное – изменения в структуре гена вызванных мутацией, ведет к изменению признака.
Синтез белка в клетке
Главным вопросом генетики является вопрос о синтезе белка. Обобщив данные по строению и синтезу ДНК и РНК, Крик в 1960г. предложил матричную теорию синтеза белков, основанную на 3–х положениях:
1. Комплементарность азотистых оснований ДНК и РНК.
2. Линейная последовательность расположения генов в молекуле ДНК.
3. Передача наследственной информации может происходить только с нуклеиновой кислоты на нуклеиновую или на белок.
С белка на белок передача наследственной информации невозможна. Таким образом матрицей для синтеза белка могут быть только нуклеиновые кислоты.
Для синтеза белка необходимы:
1. ДНК (гены) на которых синтезируются молекулы.
2. РНК – (и-РНК) или (м-РНК), р-РНК, т-РНК
В процессе синтеза белка различают этапы: транскрипции и трансляции.
Транскрипция – перепись (переписывание) информации о нуклеиновом строении с ДНК на РНК (т-РНК, и РНК, р-РНК).
Считывание наследственной информации начинается с определенного участка ДНК, который называется промотором. Промотор расположен перед геном и включает около 80 нуклеотидов.
На наружной цепи молекулы ДНК синтезируется и-РНК (промежуточная) служащая матрицей для синтеза белков и поэтому называется матричной. Она является точной копией последовательности нуклеотидов на цепи ДНК.
В ДНК имеются участки, которые не содержат генетической информации (интроны). Участки ДНК содержащие информацию называются экзонами.
В ядре имеются специальные ферменты, вырезающие интроны, а фрагменты экзона «сращиваются» между собой в строгом порядке в общую нить, этот процесс называется «сплайсингом». В процессе сплайсинга образуется зрелая м-РНК, содержащая информацию, необходимую для синтеза белка. Зрелая и-РНК (матричная РНК) проходит через поры ядерной мембраны и поступает в каналы эндоплазматической сети (цитоплазму) и здесь соединяется с рибосомами.
Трансляция – последовательность расположения нуклеотидов в и-РНК, переводится в строго упорядоченную последовательность расположения аминокислот в молекуле синтезируемого белка.
Процесс трансляции включает 2 этапа: активирование аминокислот и непосредственно синтез белковой молекулы.
Одна молекула м-РНК соединяется с 5-6 рибосомами, образуя полисомы. Синтез белка происходит на молекуле м-РНК, причем рибосомы продвигаются вдоль нее. В этот период находящиеся в цитоплазме аминокислоты активируются специальными ферментами, выделяемыми ферментами, выделяемыми митохондриями причем каждая из них своим специфическим ферментом.
Почти моментально аминокислоты связываются с другим видом РНК – низкомолекулярной растворимой, выполняющий функции переносчика аминокислот к молекуле м-РНК и получившей название транспортной (т-РНК). т-РНК переносит аминокислоты к рибосомам на определенное место, где к этому времени оказывается молекула м-РНК. Затем аминокислоты соединяются между собой пептидными связями и образуется белковая молекула. К концу синтеза белка молекула постепенно сходит с м-РНК.
На одной молекуле м-РНК образуется 10-20 молекул белка, а в некоторых случаях и много больше.
Наиболее неясным в синтезе белков вопрос о том, как т-РНК находит соответствующий участок м-РНК, к которому должна быть присоединена приносимая ею аминокислота.
Последовательность расположения азотистых оснований в ДНК, определяющая размещение аминокислот в синтезируемом белке – генетический код.
Поскольку одна и та же наследственная информация «записана» в нуклеиновых кислотах четырьмя знаками (азотистыми основаниями), а в белках – двадцатью (аминокислотами). Проблема генетического кода сводится к установлению соответствия между ними. Большую роль в расшифровке генетического кода сыграли генетики, физики, химики.
Для расшифровки генетического кода прежде всего необходимо было выяснить какое минимальное число нуклеотидов может определять (кодировать) образование одной аминокислоты. Если бы каждая из 20 аминокислот кодировалась одним основанием, то ДНК должна была бы иметь 20 различных оснований, фактически же их только 4. Очевидно, сочетание двух нуклеотидов также недостаточно для кодирования 20 аминокислот. Оно может кодировать лишь 16 аминокислот 42 = 16.
Тогда было предложено, что код включает 3 нуклеотида 43 = 64 комбинации и следовательно, способно кодировать более чем достаточное число аминокислот для образования любых белков. Такое сочетание трех нуклеотидов называется триплетным кодом.
Код имеет следующие свойства:
1.Генетический код триплетный (каждая аминокислота кодируется тремя нуклеотидами).
2.Вырожденность – одна аминокислота может кодироваться несколькими триплетами, исключение составляет триптофан и метионин.
3.В кодонах для одной аминокислоты первые два нуклеотида одинаковы, а третий изменяется.
4.Неперекрывающийся – триплеты не перекрывают друг друга. Один триплет не может входить в состав другого, каждый из них самостоятельно кодирует свою аминокислоту. Поэтому в полипептидной цепи рядом могут находиться любые две аминокислоты и возможны какие угодно их сочетания, т.е. в последовательности оснований ABCDEFGHI, первые три основания кодируют 1 аминокислоту (ABC-1), (DEF-2) и т.д.
5.Универсален, т.е. у всех организмов для определенных аминокислот кодоны одинаковы (от ромашки до человека). Универсальность кода свидетельствует о единстве жизни на земле.
6.Коленеарность – совпадение расположения кодонов в и-РНК с порядком расположения аминокислот в синтезирующийся полипептидной цепи.
Кодон – триплет нуклеотидов, кодирующий 1 аминокислоту.
 АУЦ
АУЦ
кодоны
 ЦГГ
ЦГГ
7.Бессмысленный – он не кодирует никакой аминокислоты. Синтез белка на этом месте прерывается.
В последние годы выяснилось, что в митохондриях нарушается универсальность генетического кода, четыре кодона в митохондриях изменили свой смысл, например, кодон УГА – отвечает триптофану вместо «СТОП» - прекращение синтеза белка. АУА – соответствует метионину – вместо «изолейцина».
Открытие новых кодонов у митохондрий может служить доказательством того, что код эволюционировал, и что он не сразу стал таким.
Пусть наследственной информации от гена к молекуле белка можно выразить схематически.
ДНК – РНК – белок
Изучение химического состава клеток показал, что различные ткани одного и того же организма содержат различный набор белковых молекул, хотя они имеют и одинаковое количество хромосом, и одинаковую генетическую наследственную информацию.
Отметим такое обстоятельство: несмотря на наличие в каждой клетке всех генов целого организма, в отдельной клетке работают очень немногие гены – от десятых долей до нескольких процентов от общего числа. Остальные же участки «молчат», они заблокированы специальными белками. Это и понятно, зачем, например, генам гемоглобина работать в нервной клетке? То как клетка диктует, каким генам молчать, а каким работать, следует предполагать, что в клетке имеется какой-то совершенный механизм, регулирующий активность генов определяющий, какие гены в данный момент должны быть активными и каким следует находиться в неактивном (репрессивном) состоянии. Такой механизм по данным французских ученых Ф. Жакобо и Ж. Моно получил название индукции и репрессии.
Индукция – возбуждение белкового синтеза.
Репрессия – подавление белкового синтеза.
Индукция обеспечивает работу тех генов, которые синтезируют белок или фермент, и который необходим на данном этапе жизнедеятельности клетки.
У животных важную роль в процессе регуляции генов играют гормоны клеточные мембраны; у растений – условия внешней среды и другие высокоспециализированные индукторы.
Пример: при добавлении гормона щитовидной железы в среду совершается быстрое превращение головастиков в лягушек.
Для нормальной жизнедеятельности бактерии Е (Coli) необходим молочный сахар (лактоза). Если среда, в которой находятся бактерии, лактозы не содержит, эти гены находятся в репрессивном состоянии (т.е. они не функционируют). Внесенная в среду лактоза является индуктором, включающим в работу гены, отвечающих за синтез ферментов. После удаления лактозы из среды синтез этих ферментов прекращается. Таким образом, роль репрессора может выполнять вещество, которое синтезируется в клетке, и если его содержание превышает норму или оно израсходовано.
В синтезе белка или ферментов участвуют различные типы генов.
Все гены находятся в молекуле ДНК.
По своим функциям они не одинаковы:
- структурные – гены, влияющие на синтез какого-то фермента или белка, расположены в молекуле ДНК последовательно друг за другом в порядке их влияния на ход реакции синтеза или еще можно сказать структурные гены – это гены, которые несут информацию о последовательности аминокислот.
- акцепторные – гены не несут наследственной информации о строении белка, они регулируют работу структурных генов.
Перед группой структурных генов расположен общий для них ген – оператор, а перед ним – промотор. В целом эта функциональная группа называется опереном.
Вся группа генов одного оперона включается в процесс синтеза и выключается из него одновременно. Включение и выключение структурных генов составляет сущность всего процесса регуляции.
Функцию включения и выключения выполняет особый участок молекулы ДНК – ген оператор. Ген оператор является начальной точкой синтеза белка или как говорят «считывания» генетической информации. дальше в той же молекуле на некотором расстоянии расположен ген – регулятор, под контролем которого вырабатывается белок называемый репрессором.
Из всего сказанного видно, что синтез белка происходит очень сложно. Генетическая система клетки, используя механизмы репрессии и индукции, может принимать сигналы о необходимости начала и окончания синтеза того или иного фермента и осуществлять этот процесс с заданной скоростью.
Проблема регуляции действия генов у высших организмов имеет большое практическое значение в животноводстве и медицине. Установление факторов, регулирующих синтез белка, раскрыло бы широкие возможности управления онтогенезом, создания высокопродуктивных животных, а также устойчивых животных к наследственным заболеваниям.
Контрольные вопросы:
1.Назовите свойства генов.
2.Что такое ген?
3.Назовите каково биологическое значение ДНК, РНК.
4.Назовите этапы синтеза белка
5.Перечислите свойства генетического кода.
ЛЕКЦИЯ № 7
Генетика микроорганизмов
1. Строение генетического материала у бактерий и вирусов.
2.Передача наследственности у бактерий и вирусов на примере (трансформации, трансдукции и коньюгации).
Вплоть до 30-х годов фактически все генетические исследования проводились с растениями и животными.
С изобретением в конце ХУП века микроскопа Антон ванн Левенгуком были открыты бактерии среди неведомого до тех пор мира живых форм неразличимых невооруженным глазом.
Одноклеточные организмы в том числе бактерии и вирусы отличаются от высших организмов меньшей защищенностью наследственного материала от внешней среды, что влечет за собой изменчивость частым возникновением мутаций.
Жизненный цикл бактерий длится меньше часа, каждые 20-45 минут и миллиарды которых можно вырастить за ночь в крохотном объеме питательной среды.
Все это сделало бактерии и вирусы объектом исследований. К классическим объектам генетических исследований среди бактерий относится кишечная палочка, бактерии рода сальмонелла, нейроспора, а среди вирусов – бактериофаги поражающие эти бактерии и вирус табачной мозаики.
Бактерии имеют огромное значение для человека. Многие отрасли сельского хозяйства, а также промышленности зависят от деятельности бактерий. Такие важные химические соединения, как этиловый и бутиловый спирт, уксусная кислота, ацетон образуется за счет деятельности бактерий.
Бактерии используются при производстве кефира, сыра при производстве лекарственных препаратов при биологической очистке воды загрязненных источников и т.д.
Однако при всех положительных качествах, очень многие бактерии приносят человеку животным и растениям вред, т.е. они являются возбудителями различных заболеваний: дифтерии, дизентерии, брюшного тифа и др.
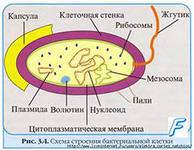
Бактерии имеют очень мелкие размеры от 1 до 10 микрометра. Форма их различна. Снаружи окружена плотной оболочкой. В отличии от многоклеточных микроорганизмов у бактерии нет четко оформленного ядра. Его заменяют по форме и величине структуры, носящие название ядерной вакуоли или нуклеоида.
Нуклеоид – ядро у прокариот. Он состоит из замкнутой в кольцо двухспиральной нити ДНК длиной 1,1-1,6 нм, которую рассматривают как одиночную бактериальную хромосому.
Цитоплозма – содержимое бактериальной клетки, отделенное цитоплазмсатической мембраной. Состоит из рибосом, внутриплазматических мембран, включений.
Рибосомы – органоиды осуществляющие синтез белка. Состоят из белка и РНК, соединенных в комплекс водородными связями. Одна бактериальная клетка содержит 5000-50000 рибосом, посредством и-РНК, они соединяются в полисомы, состоящие из 50-55 рибосом.
В цитоплазме также содержатся внехромосомные наследственные факторы, названные плазмиды.
ДНК бактерий не отличается по своему строению от ДНК высших организмов, т.е. содержат те же 4 основания: аденин, тимин, гуанин, цитозин присоединенные к дезоксирибозе. Молекулы ДНК бактерий находятся в основном в нуклеоидах и окружены хроматином, напоминая хромосомы высших организмов, виде замкнутой в кольцо 2-х спиральной нити, плотной уложенной. Один конец ДНК прикреплен к клеточной мембране.
Репликация ДНК микроорганизмов происходит также как и у высших организмов. В репликации участвуют ферменты ДНК-полимеразы.
Синтез (репликация) ДНК бактерии начинается еще в период роста, всегда с одной определенной точки и дет в одном направлении.
Деление нуклеоида заканчивается до деления клетки. В ДНК бактерии имеются отдельные участки, отвечающие за синтез определенных ферментов – называемых цистронами у высших организмов – генами.
Цистроны расположены в молекуле ДНК в линейном порядке в последовательности их влияния на синтез определенного фермента.
В период репликации ДНК и образования перегородки клетка непрерывно растет, идет формирование рибосом и других соединении.
На определенной стадии дочерние клетки отделяются др. от друга и каждая дочерняя бактериальная клетка имеет такой же набор генетической информации – какой был в исходной бактериальной клетки.
Строение вирусов. Вирусы относятся к микроорганизмам хотя резко отличаются от всех известных клеточных форм жизни. Вирусные частицы подобно бактериям не одинаковы по форме и очень малы (20-40 нм). Так вирус фага сени Т (Т1,Т2,Т4 и т.д.) кишечной палочки по форме напоминают сперматозоиды и состоят из головки и хвоста, а вирус табачной мозаики – палочковидной формы, как початок кукурузы. Для вирусов характерно более простое строение, чем у бактерий.

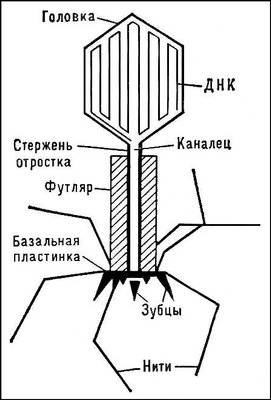
У вирусов имеется белковый чехол, который покрывает головку и хвостовую нить (отросток). В головке плотно упакована ДНК. Конец хвоста фага морфологически довольно слажен. Им фаг прикрепляется к поверхности поражаемой клетки. В зависимости от вида организма вирусные частицы содержат только ДНК или только РНК. Вирусы растений содержат РНК. Среди вирусов животных и человека существуют виды, содержащие как ДНК, так и виды с РНК. У большинства вирусов ДНК имеет двухцепочное строение, но сеть вирусы с одноцепочной ДНК.Вирусы паразитирующие в бактериях называются фагами или бактериофагами. Всюду где размножаются бактерии обнаруживаются и паразитирующие в них фаги. Они находятся в кишечнике человека, в сточных водах, почве и т.д.
Вне клетки вирусные частицы инертны и не размножаются. Проникают вирусы в клетку бактерии или высшего организма при механическом повреждении тканей организма или путем прикрепления к поверхности клетки концевыми нитями отростков растворяя ее оболочку специальным ферментом впрыскивая в клетку частицы ДНК или РНК с незначительным проникновением белка. После проникновения в клетку хозяина ДНК вируса начинает интенсивный синтез, образуя новые молекулы ДНК. И под их контролем синтезируются специфические для данного вируса белки.
Для белкового синтеза используются рибосомы хозяина. В это время нормальные биологические процессы и синтез белков «своих» в клетке хозяина прекращаются и все происходящие в ней процессы подчиняются деятельности ферментов. В результате в клетке образуется большое количество вирусного белка, молекул ДНК или РНК вируса, что заканчивается распадом клетки хозяина ее лизисом.
Синтез вирусного белка начинается через несколько минут после заражения организма хозяина. Одна частица фага, попавшая в клетку, способна дать начало сотне и более новых частиц. Через 16-18 минут происходит лизис. После разрушения, или (лизиса) клетки, эти частицы входят в окружающую среду вполне зрелыми (готовыми) к новому заражению. В зависимости от взаимодействия вирусов с клеткой хозяина они подразделяются на вирулентные и умеренные.
Фаг, способный вызвать лизис клетки – называется вирулентным. Умеренные фаги проникают в клетку хозяина не разрушают ее, а остаются в ней, в качестве жильцов, т.е. вступают в симбиоз. Бактерии несущие умеренный фаг – называются лизогенными, а сам фаг, существующий в подобной форме – профагом. Профаг может существовать с бактериальной клеткой длительное время, но может переходить и в лизис. Экспериментами установлено, что участок ДНК умеренного фага всегда прикрепляется к соответствующему участку молекулы ДНК бактерии. Синтез ДНК профага происходит синхронно с репликацией ДНК бактерии. Основная форма размножения у бактерии – деление клеток, у вирусов – редупликация ДНК, РНК и образование новых частиц в клетке хозяина.
Однако между клетками разных бактерий или частицами вируса происходит обмен генетическим материалом, осуществляемый у вирусов – путем трансформации, трансдукции, коньюгации.
Трансформация у бактерий и вирусов
Трансформация – изменение наследственности бактериальной клетки путем проникновения в нее ДНК, происходящей из другой клетки или штамма с др. наследственными задатками.
В процессе трансформации участвуют 2 бактериальные клетки: донор и реципиент причем они не соприкасаются друг с другом.
Механизм переноса генетического материала заключается в том, что из клеток донора выделяются в окружающую среду ДНК.
Явление трансформации было открыто в 1928 Гриффитсом. Получив у пневмококков, вызывающих заболевание мышей воспаление легких два штамма – один капсульный вирулентный (ядовитый) гладкий S – формы образует массивную полисахаридную капсулу, придает колонии пневмококков блестящий вид. Капсула защищает бактерию от разрушающего действия обычных защитных механизмов тканей инфицированного животного. Второй – бескапсульный, авирулентный (не ядовитый). R- формы такой капсулы не образует. Он вводил мышам:
Если штамм был вирулентным – мыши заболевали и погибали;
Если он был авирулентным R – оставались здоровыми.
При введении мышам убитого нагреванием вирулентного штамма они заболевали, когда тем же мышам был введен вслед за ним второй – авирулентный штамм, у них развивалось воспаление легких и они погибали.
Штамм, выделенный из погибших мышей оказался вирулентным, но бескапсульным.
Таким образом, в организме мышей произошла передача вирулентности от одного штамма бактерий другому. Это явление было названо трансформацией.
В 1944 году Эйвери с сотрудниками повторил эксперимент Гриффитса. Трансформация происходит не часто – примерно 1 на 1000 клеток, а межвидовая – еще реже (1 на 3-10 млн.клеток).
У вирусов трансформация является единственным путем обмена генетическим материалом между разными штаммами.
Трансдукция
В 1952 году публикуются работы Н.Д.Циндера и Дж.Ледерберга в которых описано явление трансдукции, также подтверждающие решающую роль ДНК в передаче наследственной информации. Они проводили исследования на патогенных для мышей бактериях Salmonellatyphimurium
Суть трансдукции состоит в том, что бактериофаги (паразиты бактерий) из одной бактериальной клетки в другую вместе со своей ДНК переносят отдельные гены. Трансдуцированные бактерии приобретают только те свойства, которые были у донора.
Опыт позволивший открыть этот новый генетический механизм и новый способ изучения наследственности заключается в следующем.
Были отобраны два штамма этих бактерий: штамм 22А ауксотрофный, не способный синтезировать триптофан (Т~), и штамм 2А, способный синтезировать триптофан (Г1"). Эти штаммы засевали в U-образную трубку, разделенную внизу бактериальным фильтром. В одно колено трубки засевали штамм 22А (Т~), в другое — штамм 2А (Т4"). После определенного периода инкубации бактерии штамма 22А при посеве на минимальную питательную среду дали небольшое количество колоний (частота появления трансдуцированных клеток была равна 1-10—*). Это свидетельствовало о том, что некоторые клетки приобрели способность синтезировать триптофан. Каким же образом бактерии могли приобрести это свойство?
Исследования показали, что штамм 22А был лизогенен по фагу Р-22. Этот фаг освобождался из лизоген-ной культуры, проходил через фильтр и лизировал штамм 2А. Присоединив часть генетического материала штамма 2А, фаг возвращался обратно и передавал этот генетический материал штамму 22А. Штамм 22А приобретал специфические наследственные свойства штамма 2А, в данном случае свойство синтезировать триптофан. Аналогичным образом могут быть трансдуцированы и другие признаки, в том числе способность к сбраживанию, устойчивость к антибиотикам и т. д. Существуют три типа трансдукции: неспецифическая (общая), специфическая и абортивная.
Общая (неспецифическая) трансдукция - перенос бактериофагом фрагмента любой части бактериальной хромосомы. В клетке, инфицированной бактериофагом, в ходе сборки дочерней популяции в головки некоторых фагов может проникнуть фрагмент бактериальной ДНК или плазмиды либо вместе с вирусной ДНК, либо вместо нее. Этот процесс происходит вследствие того, что бактериальная ДНК фрагментируется после фаговой инфекции и кусочек бактериальной ДНК того же размера, что и фаговая ДНК, проникает в вирусную частицу с частотой приблизительно 1 на 1000 фаговых частиц. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
Специфическая трансдукция наблюдается в том случае, когда фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы в результате случайного процесса захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Так как большинство умеренных фагов интегрируют в бактериальную ДНК в специфических участках, для таких бактериофагов характерен перенос в клетку-реципиент определенного участка бактериальной ДНК донора. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.Наиболее характерным примером служит трансдукция, осуществляемая фагом λ. Обычно он трансдуцирует определенные гены: gal (кодирует синтез галактозы) и bio (кодирует синтез биотина). При переходе в состояние профага фаг λ включается в определенный участок хромосомы бактерии-хозяина - между генами gal и bio. Отделение фаговой ДНК от бактериальной хромосомы может происходить неточно и какой-то фрагмент ее останется в хромосоме, а близко расположенные гены будут захвачены фаговой ДНК. В случае заражения трансдуцирующим фагом клеток, дефектных по определенному гену, например gal -, может произойти рекомбинация с заменой собственного дефектного гена бактерии интактным трансдуцированным геном с образованием рекомбинанта (трансдуктанта) gal +.
Абортивная трансдукция. При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Коньюгация у бактерий
Коньюгацией называется передача генетического материала т одних бактерий другим при скрещивании.
Явление коньюгации бактерии открыто в 1946 году Дж.Ледербергом и Е.Татумом (на щтамме К-12 кишечная палочка).
Коньюгация у бактерий представляет собой подобие примитивного полового процесса. При этом одни клетки являются «мужскими» (служат донорами генетического материала, а другие – «женскими» играют роль реципиентов.) 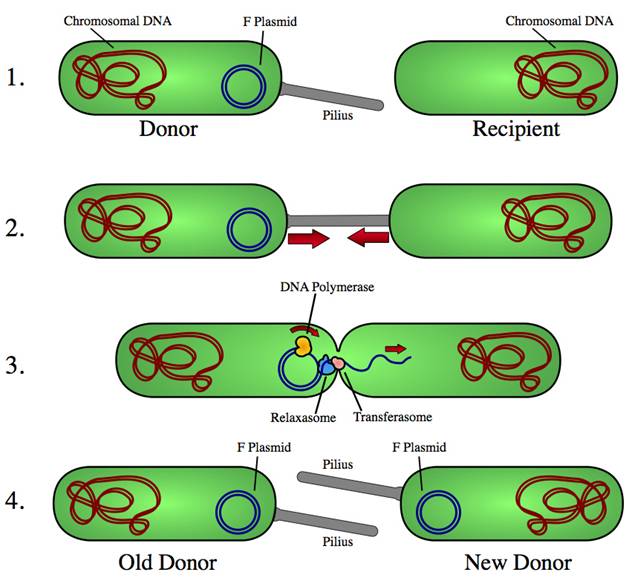
При коньюгации бактерии сближаются, клеточная оболочка в точке их соприкосновения растворяется и между ними образуется цитоплазматический мостик, по которому хромосома одного штамма переходит в другой.
У.Хейс (1953г.) показал, что генетический материал передается односторонне, всегда от клеток одного определенного штамма к другому, обратного процесса не наблюдается (т.е. от «мужской» клетки к «женской»).
Затем Э.Вольман и Ф.Жакоб (1962 г.) установили, что этот процесс строго ориентированный, т.е. хромосома (генетический материал) донора внедряется в «женскую клетку» лишь одним определенным концом. Причем продвигается очень медленно, в минуту проходит 1/160 хромосомы. Вся нить ДНК из клетки в клетку переходит за 2 часа.
Так как цитоплазматический мостик – образование очень хрупкое, то даже при небольшом встряхивании он разрушается. Вот почему целая хромосома донора переходит в клетку реципиента очень редко. О величине перешедшего в бактерию реципиента участка молекулы ДНК донора можно судить по тем рекомбинациям, которые возникают у клетки реципиента.
Таким образом, процесс коньюгации дает возможность изучить расположение генов в хромосомах бактерии.
О расстоянии между генами в картах хромосом судят по тому времени, которое прошло с начала коньюгации, и о порядке расположения генов в хромосоме.- по последовательности их перехода в клетку реципиента. В результате исследований составлены карты хромосом.
ЛЕКЦИЯ № 8
Мутационная изменчивость
1. Мутационная теория Де-Фриза
2. Классификация мутаций
3. Факторы, вызывающие мутагенез
4. Репарирующие системы клеток
5. Защита животных от мутагенов
6. Закон гомологических рядов академика Н.И. Вавилова

