




Из всех областей вопросов, относящихся к компетенции общей физиологии, ни одна не является столь специфически человеческой, как область физиологии двигательных функций, несмотря на наличие здесь бесспорной и непрерывной преемственности от филогенетических предков. Дело в том, что больше ни в одной системе физиологических функций не имел места такой интенсивный и вдобавок убыстряющийся филогенетический прогресс. Едва ли мы смогли бы отметить существенные сдвиги в смысле эволюционного прогресса между любым представителем теплокровных и человеком в какой бы то ни было иной функциональной сфере. Исключение в смысле, несомненно, еще более бурного прогресса составляет только область, выделенная по методологическому признаку в ведение самостоятельной науки - область явлений психической жизни, или область центральных замыкательных систем нервного аппарата [1].
В процессе эволюции соматической системы (разве лишь за исключением самого последнего филогенетического отрезка) определяющим звеном являются эффекторные функции. Судьбу индивидуума в борьбе за существование решают его действия - большая или меньшая степень их адекватности во все осложняющемся процессе приспособления. Рецепторика здесь представляет собой уже подсобную, обслуживающую функцию. Нигде в филогенезе созерцание мира не фигурирует как самоцель, как нечто самодовлеющее. Рецепторные системы являются либо сигнальными, либо они процессуально обеспечивают полноценную, координированную работу эффекторов. Таким образом, и в сигнальной, и в коррекционной роли рецепторы состоят при эффекторных аппаратах, влияя на биологическую судьбу особи или вида не иначе, как через эти последние. Центральные замыкательные системы в этом аспекте исторически являются уже подсобными приборами для подсобных [1,2].
Усложнение возникающих перед организмом двигательных задач и откликающееся на него обогащение координационных ресурсов особи совершаются по двум линиям. С одной стороны, двигательные задачи делаются более сложными в прямом смысле слова. Возрастает разнообразие реакций, требующихся от организма. К самим этим реакциям предъявляются более высокие требования в отношении дифференцированности и точности; наконец, осложняется смысловая сторона движений, действий и поступков животного. Достаточно напомнить, насколько, например, аэродинамический полет птицы сложнее почти полностью гидростатического плавания рыбы или насколько богаче по контингентам участвующих движений охота хищного млекопитающего по сравнению с охотой акулы. Молодая отрасль проворных теплокровных млекопитающих победила тугоподвижных юрских завров именно своею более совершенной моторикой. С другой стороны, в общем составе встающих перед организмом двигательных задач все возрастает процент задач разовых, непредвиденных, экстемпоральных за счет более древних шаблонных ситуаций. Все многочисленные исследования "пластичности нервной системы" показывают наряду с эволюционным возрастанием приспособляемости центральной нервной системы к нетрафаретным изменениям условий немедленность, почти мгновенность ее перестроек при самых фантастических постановках опыта. Но даже если оставить в стороне эксквизитные экспериментальные анастомозы мышц и нервов, то гораздо более будничный факт возрастающей по ходу филогенеза способности к накоплению индивидуального опыта, к замыканию новых условных связей, т.е. опять-таки к выходу за рамки родовых стереотипов, подтверждает высказанное положение [2].
Слегка схематизируя, можно сказать, что первая из двух упомянутых линий развития двигательных координации обеспечивается и сопутствуется преимущественно эволюцией рецепторики, вторая - эволюцией центральных замыкательных систем.
Ход филогенетического развития строения центральной нервной системы, в отличие от всех прочих органов и систем тела, состоит не только (и даже не столько) в количественном разрастании, сколько в качественном обрастании ее новыми образованиями, не имеющими гомологов в предшествующих этапах филогенеза и по большей части представляющими собой надстройки на один (или больше) нейрональный этаж на пути следования нервного процесса. Этот принцип приводит к неминуемой скачкообразности развития центральной нервной системы уже из-за дискретности нейронной схемы: осложнение рефлекторной дуги или вообще любого маршрута нервного импульса внутри центральной нервной системы возможно не иначе, как на целое число новых промежуточных нейронов.
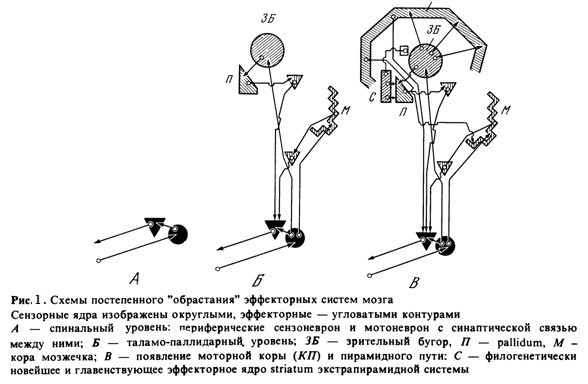
Путь, по которому центральная нервная система в своем развитии преодолевает эту скачкообразность, вскрывается эпизодами, подобными, например, ходу развития зрительного аппарата от амфибий до птиц, с переходом его от однонейронной схемы "сетчатка - ядра покрышки" к двухнейронной: "сетчатка - наружные коленчатые тела - зрительная зона коры полушарий». На протяжении какого-то этапа развития оба аппарата, и старый, и новый, действуют рядом, после чего первый или инволюционирует, как это и случилось с мезэнфалическими центрами зрения, или, чаще, модифицируется так, чтобы образовать вместе со вторым более сложный функциональный синтез. Так было, например, с постепенным развитием эффекторных аппаратов мозга (рис. 1) [22].
Усложнение двигательных задач, неминуемо требующих разрешения со стороны особи, и само по себе совершается отнюдь не плавно и постепенно; наоборот, перемены в образе жизни, зоологическом окружении, экологической обстановке и т.д. приводят к накоплению все больших масс качественно новых координационных проблем с не встречавшимися ранее и не имевшими возможности войти в обиход особыми чертами смысловой структуры, двигательного состава и т.д. В течение какого-то времени животные справляются с этими необычными задачами при помощи своих наличных ресурсов; однако рано или поздно противоречие между новыми смысловыми и сензорными качествами нахлынувших задач и неадекватными им координационными средствами животного приводит путем отбора к преобладанию особей, способных справиться с этими новыми качествами, и этим сразу, скачком, получить в свое распоряжение целый новый класс движений, однородных по своему типу и уровню сложности и сходных между собой по качествам потребного сензорного контроля. Если бы эволюционное развитие совершалось по Ламарку, в порядке постепенного упражнения рабочих органов, то можно было бы, пожалуй, ожидать каких-либо гипертрофических, количественных, постепенно образующихся приспособительных изменений мозга. Но, осуществляясь по принципу отбора, развитие центральной нервной системы в ответ на новые классы двигательных задач не может протекать иначе, как в виде накапливающегося преобладания индивидуумов с качественно отличным, мутировавшим в каких- то отношениях мозгом. Возникновение в филогенезе очередной новой мозговой надстройки знаменует собой биологический отклик на новое качество или класс двигательных задач. Как будет показано ниже, это обязательно означает в то же время появление нового синтетического сензорного поля, а тем самым и появление возможности реализации нового класса или контингента движений, качественно иначе строящихся и иначе управляемых, нежели те, которые были доступны виду до этих пор. Мы обозначаем всю перечисленную совокупность морфологических и функциональных сторон, характерных для такого нового класса движений, как очередной уровень построения движений и двигательных координаций [1,2,21].
Сказанное выше о линиях усложнения двигательных задач, возникающих перед организмом, позволяет оценить и те направления, по которым совершалось поочередное развитие возникавших один за другим координационных уровней построения.
Более новые в филогенезе, они же и более высокие, уровни становятся:
1) все более тесно связанными с телерецепторикой и надстроенными над ней обобщающими системами в коре головного мозга;
2) все более экстемпоральными, т.е. пригодными для осуществления разовых координационных решений и пластических переключений;
3) все более синтетичными, т.е. опирающимися на сложные психологически организованные синтетические сензорные поля;
4) все более богатыми мнестическими элементами, накопленными из индивидуального опыта. В этих же направлениях изменяются и облики тех движений, и действий, которые ведутся на соответственных уровнях.
Каждый новый уровень приносит с собой комплект новых движений, какие раньше были организму недоступны. Следует сразу отвергнуть как неверное, старое представление, будто филогенетически более молодые надстройки обеспечивают в основном новые качества координации и, следовательно, будто каждый из разновозрастных мозговых морфологических этажей равнозначен какой-то одной стороне координационной отделки любого целостного движения. Каждый новый морфологический этаж мозга, каждый очередной функциональный уровень построения содержит и приносит с собой не новые качества движений, а новые полноценные движения. В нервной системе высокоразвитого позвоночного содержащимся в ней N структурным этажам и доступным для нее N уровням соответствует N особых списков или контингентов движений, вполне законченных и биологически пригодных для решения определенных, посильных им задач. Было бы очень трудно понять, какой биологический смысл и какое оправдание своего существования могли бы иметь движения-недоноски, лишенные в течение долгих веков филогенетической эволюции какой-либо существенной группы координационных качеств или, наоборот, представляющие собой наборы второстепенных, вспомогательных качеств без самого главного смыслового определителя - фон без фигуры. В истории развития каждый из уровней построения, констатируемых у человека, был на каком-то этапе наивысшим (разумеется, с известными поправками в отношении эволюции контингентов) и определял собой "потолок" координационных возможностей организма, обрывавший сверху список доступных ему в ту пору движений; но на каждом подобном этапе эти движения были вполне закруглены и координационно оформлены в меру тех скромных двигательных задач, какие им предстояло разрешать [5].
Центральная нервная система высокоразвитого позвоночного, например антропоида или человека, представляет собой своего рода геологический разрез, отображающий в сосуществовании всю историю развития нервных систем, начиная от диффузных невропилей низших беспозвоночных и простейших спинальных рефлекторных дуг первобытных хордовых. Все это воспроизведено в такой высокоорганизованной нервной системе, в ее последовательных наслоениях, этажах и надстройках с не меньшей точностью, чем, например, индивидуальная история дерева - в его древесных кольцах [1].
На фоне этого факта представляется очень интересным и многозначительным, что координационные контингенты движений человека образуют точно такого же рода симультанную рекапитуляцию всей истории животных движений, начиная от таких продвижений, как перистальтикоподобные движения кольчатого червя или глотательнорвотные движения голотурии. Такая рекапитуляция обнаруживает при этом неоспоримые преимущества перед невроморфологией, поскольку воспроизводит филогенез не в статике и не в символике гистологических обликов нервных ядер, ничего не сообщающих нам о своей функциональной сущности, а в динамике, в самих движениях, доступных точным сравнениям как по своему содержанию и смыслу, так и по своему оформлению, с движениями современных нам представителей всех ступеней филогенетической лестницы. У самого дна глубокой шахты, опускаемой нами в толщу двигательных координации человека, мы находим древнейшие палеокинетические координации, отошедшие у высших позвоночных в удел вегетативным отправлениям: перистальтические движения кишечника, стрикционно- дилятационные движения в сосудистой системе, сфинктерах желудка, желчного пузыря, прямой кишки и т.п. Поднимаясь выше, мы встречаемся с первичными, самыми элементарными и по структуре, и по определяющей их афферентации неокинетическими координациями — спинальными рефлексами, подробно изученными школой Sherrington. Еще выше мы вступаем в область движений с более сложной биологической мотивировкой и с афферентацией, синтетически включающей как телерецепции, так и индивидуальные мнестические компоненты, — в область подлинной психофизиологии. Еще более кверху, еще в большей и более невозместимой мере зависящие от деятельности коры полушарий залегают самые молодые в филогенезе специфически человеческие координации, мотивы к возникновению которых уже никак нельзя свести к чисто биологической причинности: в первую очередь координации речи, письма и предметных, трудовых действий с их социально-психологической обусловленностью. Каждое из этих последовательных наслоений связано с очередным новым морфологическим субстратом, и каждое, не отрицает нижележащих, более древних координационных напластований, но сливается с ними в очень своеобразный и многообразный синтез [1,2,10].
Классификация движений
Все многообразие форм движения животных и человека основывается на физических законах перемещения тел в пространстве. При классификации движений необходимо учитывать конкретные целевые функции, которые должна выполнять двигательная система. Движение - одно из проявлений жизнедеятельности, обеспечивающее организму возможность активного взаимодействия со средой, в частности, перемещение с места на место, захват пищи и др. [6]
В самом общем виде таких функций четыре:
1 поддержание определенной позы;
2 ориентация на источник внешнего сигнала для его наилучшего восприятия;
3 перемещения тела в пространстве;
4 манипулирование внешними вещами или другими телами.
Иерархия уровней мозгового управления движениями также находится в зависимости от требований к структуре движения. Установлено, что подкорковый уровень связан с набором врожденных или автоматизированных программ [5].

