




Структура биотического сообщества слагается из нескольких показателей.
Обилие — число особей на единицу площади или объема.
Частота — отношение числа особей одного вида (n) к общей численности особей (N), выраженное в процентах (n ⋅ 100)/ N.
Постоянство — отношение числа содержащих изучаемый вид выборок (р) к общему числу выборок (Р), выраженное в процентах (р / Р) ⋅ 100. В зависимости от величины этого отношения различают следующие категории видов: постоянные — встречаются более чем в 50% выборок, добавочные — в 25-50% выборок, случайные — менее чем в 25% выборок.
Доминирование — способность вида занимать в экосистеме главенствующее положение и оказывать влияние на распределение в ней энергии. Доминанты — это те виды, которые на своем трофическом уровне облада-
178
ют наибольшей продуктивностью. Из сотен или тысяч видов, входящих в состав экосистемы, лишь немногие виды-доминанты оказывают на ее свойства определяющее влияние. Искусственное изъятие доминанта из экосистемы сказывается не только на биотическом сообществе, но и на физической среде.
Влияние, которое оказывают виды-доминанты на структуру и свойства экосистемы, обусловливается их численностью, размером особей, продукцией. Так, среди видов, особи которых крупные (например, деревья), степень доминантности определяется биомассой, а среди видов, особи которых мелкие (травянистые растения), доминантами будут наиболее обильно представленные виды. Для оценки степени доминантности в лесу используется площадь поперечного сечения ствола, а на лугу— «проективное покрытие», т. е. площадь поверхности земли, занятой растениями данного вида.
В наземных экосистемах цветковые растения обычно доминируют не только на своем трофическом уровне, но и в пределах всего сообщества, поскольку они обеспечивают убежищем подавляющее большинство организмов сообщества и, кроме того, оказывают разнообразное влияние на абиотическую среду. Консументы также могут играть регулирующую роль в целом сообществе. Там, где растения невелики по размеру, животные довольно сильно влияют на физическую среду.
Если степень доминантности определяется в конечном итоге по уровню продуктивности, то показатель доминирования (С) представляет собой иное понятие. Он вычисляется по формуле С = Σ(ni / N)2, где ni —степень доминантности каждого вида (на основании числа особей, величины биомассы, продуктивности и т. д., но чаще всего на основании числа особей — отсюда n), N — общая степень доминантности, т.е. численность особей всех видов.
Предположим, что сообщество состоит из пяти одинаково важных видов; степень доминантности каждого вида равна 2 (например, плотность составляет 2 организма на 1 м2). Тогда
179
С = 0,04+ 0,04+ 0,04+ 0,04+ 0,04 = 0,2. В другом сообществе, также состоящем из пяти видов, у одного степень доминантности равна 6, а у всех остальных—1. Тогда С = 0,36 + 0,01 + 0,01 + 0,01 + 0,01 = 0,4. Иными словами, во втором сообществе доминирование оказывается более концентрированным, т. е. меньшее число видов являются доминантными.
Доминирование отчетливее проявляется там, где абиотические факторы носят экстремальный характер: в пустыне, тундре. Иными словами, регулирующая роль в сообществах с экстремальными условиями распределена между меньшим числом видов. Так, на севере лес может на 90% состоять из одного или двух видов деревьев, в тропическом лесу, напротив, доминантами по тому же критерию могут быть более десяти видов деревьев.
Верность — выражает степень привязанности вида к экосистеме. По этому признаку различают следующие категории видов: 1) характерные, т. е. свойственные исключительно одной экологической системе, или, что случается чаще, представленные в ней гораздо обильнее, чем в других экосистемах; 2) преферентные, т. е. встречающиеся в нескольких смежных экосистемах, но предпочитающие одну из них; 3) чуждые, т. е. случайно попавшие в сообщество, к которому они не принадлежат; 4) индифферентные, т. е. способные существовать с равным успехом во многих экосистемах. Как правило, в любой экосистеме характерных видов меньше, чем преферентных или чуждых. Зато по численности особей характерные виды преобладают над преферентными и чуждыми.
Видовое разнообразие трофического уровня или сообщества в целом определяют главным образом малочисленные редкие виды. Зависимость между числом видов и их численностью, т. е. числом особей, принадлежащих одному виду, описывается вогнутой кривой (рис. 11.1). Обычно в состав сообщества входит несколько видов с высокой численностью и множество редких видов с небольшой численностью. Показатель видового
180
разнообразия Ĥ= -Z(ni //V) log (ni / N), где ni, — степень доминантности каждого вида, /V — общая степень доминантности.
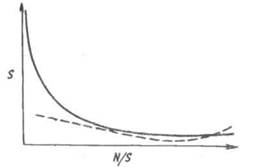
Рис. 11.1. Общая зависимость между числом видов (S) и числом особей, приходящихся на один вид (A//S).
Под влиянием стресса кривая выравнивается (пунктирная линия).
Видовое разнообразие слагается из двух компонента. Первый может быть назван видовым богатством или компонентом многообразия. Он выражается отношением S/ N — общего числа видов S кобщему числу особей N.
Второй компонент видового разнообразия — это так на 1ыиаомсШ вирппиопность распределения особей между видами. Объективным показателем служит показатель выравненности: е = Ĥ / logS.
Две экосистемы, каждая из которых состоит из 10 видов и 100 особей, могут иметь одинаковое отношение S/Л/, но разные показатели выравненности. В одном крайнем случае распределение может иметь вид: 91-1-1-1-1-1-1-1-1-1 (минимальная выравненность), а в другом — по 10 особей на вид (идеальная выравненность).
Существенное влияние на увеличение числа видов оказывает продуктивность экосистемы. В тех местообитаниях, где пищи мало, кормящиеся животные не могут позволить себе игнорировать различные типы жертвы; там же, где ее много, они могут быть более прихотливыми и
181
ограничить свою диету только наилучшими пищевыми объектами. Следовательно, в более продуктивных местообитаниях, т. е. там, где плотность пищи высока, выбор шире и поэтому возможностей для специализации больше, чем в менее продуктивных местообитаниях. Поскольку каждый вид использует только часть общего диапазона доступного корма, одинаковый спектр типов пищи в более продуктивной среде даст возможность существовать большему числу видов (рис. 11.2). Это возможно еще и потому, что в низкопродуктивных местообитаниях количество некоторых ресурсов не обеспечивает существования вида, а в продуктивных их хватает для успешной эксплуатации. Например, в открытой пустыне, где на 1 га насчитывается лишь одно муравьиное гнездо, вряд ли будет жить популяция специализированных ящериц, питающихся муравьями; вместе с тем в другой, более богатой местности, где на 1 га насчитывается несколько гнезд, ящерицы могут существовать.
Под влиянием стресса уменьшается число редких видов и возрастает роль видов, устойчивых к стрессу, т. е. происходит концентрация доминирования в этих видах. Кривая зависимости между числом видов и числом особей, принадлежащих одному виду, выравнивается (рис. 11.1). Для систем, испытавших стресс, характерна бедность видового состава, причем каждый вид многочислен.
Подобная закономерность хорошо иллюстрируется относительным обилием разных видов насекомых в водной среде, загрязненной нефтью (рис. 11.3). Если нефтяное загрязнение реки незначительное, то в зарослях высших водных растений встречается 4 вида насекомых. При сильном загрязнении остается лишь один вид — поденка Baetis vermis, устойчивая к загрязнению. В специфическом местообитании — на каменистом перекате — отмеченная взаимосвязь проявляется еще более четко.
Аналогичная картина наблюдается для агроценозов. Распашка и освоение целины в Оренбургских степях привели не только к резкому сокращению числа обитающих там видов (135 вместо 312), но и к существенному увеличению числа особей на 1 м2
182
пахотного горизонта (341 вместо 159). В наибольшей мере возросла численность серой зерновой совки и пшеничного трипса, юг да как безвредные муравьи Leptotfyorax nassonovi стали исключительно редкими. Сократилась также численность марокской саранчи и жуков-кравчиков, лишившихся привычных местообитаний на посевах пшеницы.
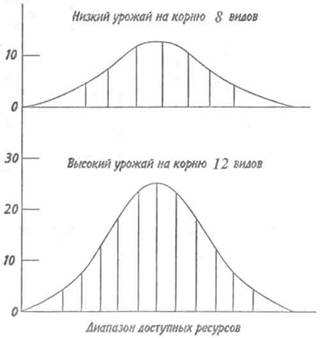
Рис. 11.2. Графическая иллюстрация возможности
увеличения числа видов при возрастании обилия
потребляемых ими ресурсов.
Горизонтальная ось — доступные ресурсы разных типов
(ранжирование любым удобным способом).
Вертикальная ось — количество ресурсов каждого типа.
Обе кривые ограничены одинаковыми отрезками горизонтальной оси, но высота нижней кривой вдвое больше высоты верхней.
Все сегменты под кривыми, кроме крайних, имеют приблизительно одинаковую площадь, которая пропорциональна одинаковому количеству ресурса. При низком урожае на корню отдельные виды должны иметь широкие ниши, и поэтому вместе может существовать только 8 видов. Если урожай на корню удвоится, ниши могут сократиться, а число видов может возрасти до 12 (по: Пианка, 1981).
183
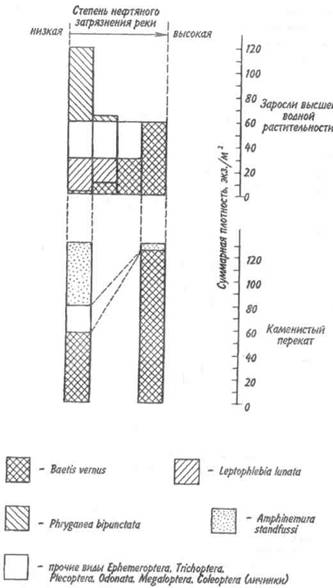
Рис. 11.3. Зависимость плотности водных насекомых
в зарослях растительности и на каменистом перекате
от степени нефтяного загрязнения реки.
184









 С концепцией видового разнообразия связан ряд важнейших экологических принципов. Большое видовое разнообразие означает более длинные пищевые цепи, больше случаев мутуализма, паразитизма, комменсализма и т.д., а также широкие возможности для действия отрицательной обратной связи, которая уменьшает колебания численности и потому повышает стабильность экосистемы. Кроме того, видовое разнообразие обеспечивает множественное дублирование пищевых цепей на уровне видов. В агроценозах такое дублирование обеспечивается на уровне особей, поэтому упрощенная маловидовая система начинает пульсировать даже при несильных стрессах.
С концепцией видового разнообразия связан ряд важнейших экологических принципов. Большое видовое разнообразие означает более длинные пищевые цепи, больше случаев мутуализма, паразитизма, комменсализма и т.д., а также широкие возможности для действия отрицательной обратной связи, которая уменьшает колебания численности и потому повышает стабильность экосистемы. Кроме того, видовое разнообразие обеспечивает множественное дублирование пищевых цепей на уровне видов. В агроценозах такое дублирование обеспечивается на уровне особей, поэтому упрощенная маловидовая система начинает пульсировать даже при несильных стрессах.
Там, где снижен расход энергии на поддержание определенной температуры тела при колебаниях значений физических факторов среды (т. е. когда отношение R/ В невелико), остается больше энергии на создание видового разнообразия. Соответственно в экосистемах со (лабильными условиями обитания, например в дождевом тропическом лесу, больше разнообразие видов.
Большое влияние на видовое разнообразие оказывают функциональные связи между трофическими уровнями. Интенсивный выпас или активное хищничество существенно снижают видовое разнообразие травостоя на пастбище или популяций жертв. В то же время умеренное «хищничество» часто снижаетплотность доминантой, давая таким образом возможность менее конкурентоспособным видам лучше использовать пространство и другие ресурсы, в результате чего видовое разнообразие возрастает.
На литорали атлантического побережья США хищная морская звезда Pisaster питается в основном мидиями, благодаря чему на субстрате постоянно открываются свободные от мидий участки, которые заселяются другими беспозвоночными, в иных условиях не выдерживающими конкуренции с мидиями. Поэтому там, где обитают морские звезды, почти вдвое больше видовое разнообразие беспозвоночных, чем там, где звезд нет. Пасущиеся копытные в степях и саваннах поддерживают определенный видовой состав и структуру травостоя,
185
а муравьи-кочевники в тропических лесах — видовой состав и численность наземных и подстилочных беспозвоночных.
Прикрепленные организмы в приливно-отливной зоне (где пространство более ограничено, чем пища) характеризуются большим видовым разнообразием в тех местах, где активно действуют первичные и вторичные хищники. В таких ситуациях экспериментальное удаление хищников снижает разнообразие всех сидячих организмов независимо от того, питались ими хищники или нет.

