




Открытие И.П. Павловым анализаторов и создание учения об условных рефлексах, в основе которого лежал объективный анализ динамики нервных процессов, послужило основой для развития современных материалистических представлений о динамической локализации мозговых функций - целостном и одновременно дифференцированном вовлечении мозга в любую из форм его активности.
Предложенный И.П. Павловым объективный условно рефлекторный метод исследования позволил наиболее адекватно подойти к экспериментальному решению проблемы функциональной организации мозга. И.П. Павлов развил и экспериментально обосновал представления об анализаторных системах, где каждый анализатор есть определенная анатомически локализованная структура от периферических рецепторных образований до проекционных зон коры головного мозга. Он предположил, что кроме локальных проекционных зон коры, выступающих в качестве «ядра коркового конца анализатора» (или проекционных зон коры), существуют периферические зоны представительства каждого анализатора, так называемые «зоны рассеянных элементов». В силу такой структурной организации все анализаторы, включая и двигательный анализатор, своими периферическими (корковыми) зонами перекрываются и образуют вторичные проекционные зоны коры, которые И.П. Павлов уже тогда рассматривал как «ассоциативный центры» мозга, составляющие основу для динамического взаимодействия всех анализаторных систем.
Даже относительно простое произвольное движение включает целый комплекс как чувствительных (афферентных), так и двигательных (эфферентных) импульсов. Произвольные движения, ходьба, или такой сложный психический процесс, как чтение, осуществляются за счет совместной деятельности нервных центров, объединенных в сложную функциональную систему по П.К. Анохину. Естественно, что сложная функциональная система не может быть «локализована» в определенном участке нервной системы.
Принципы функциональной организации человеческого мозга были разработаны видным отечественным ученым П. К. Анохиным. Они состоят в следующем. Головной мозг человека, обеспечивающий прием и переработку информации, создание программ собственных действий и контроль за их выполнением, всегда работает как единое целое.
А. Р. Лурия предложил структурно-функциональную модель мозга как субстрата психической деятельности. Эта модель характеризует наиболее общие закономерности работы мозга как единого целого и позволяет объяснить его интегративную функцию по Е. Д. Хомской. Согласно этой модели, весь мозг можно разделить на три структурно-функциональных блока:
а) энергетический блок,
б) блок приема, переработки и хранения экстероцептивной информации,
в) блок программирования, регуляции и контроля сложных форм деятельности.
Любая ВПФ осуществляется при обязательном участии всех трех блоков. Каждый блок характеризуется особенностями строения, физиологическими принципами, лежащими в основе его работы, и той ролью, которую он играет в обеспечении психических функций [Лурия А. Р.Основы нейропсихологии. М., 1973].
Первый блок — это блок регуляции энергетического тонуса и бодрствования. Было доказано И. П. Павловым, А. Р. Лурия, М. Н. Ливановым, что для нормальной психической деятельности организм должен находиться в состоянии бодрствования, т.е. кора больших полушарий должна находиться в состоянии тонуса, иметь необходимый уровень возбуждения. Только в условиях оптимального бодрствования человек может наилучшим образом принимать и перерабатывать информацию, вызывать в памяти нужные системы связей, программировать деятельность, осуществлять контроль над ней. Было установлено, что аппараты, обеспечивающие и регулирующие тонус коры, находятся не в самой коре, а в лежащих ниже стволовых и корковых отделах мозга. Таким аппаратом являются неспецифические структуры разных уровней — ретикулярная формация ствола мозга, неспецифические структуры среднего мозга, лимбическая система, область гиппокампа. Ретикулярная формация представляет собой нервную сеть, в которую вкраплены тела нейронов с короткими аксонами.
Механизм работы первого блока следующий. Постоянный тонус коры поддерживается благодаря двум источникам. С одной стороны, для сохранения бодрствующего состояния коры нужен постоянный приток информации из внешнего мира: животное, лишенное притока внешних раздражении, засыпает. Известно также, что «информационный голод» в результате длительного одиночного пребывания человека в темной и звуконепроницаемой камере вызывает галлюцинации, которые частично компенсируют недостаток в постоянном притоке внешних раздражении. Постоянный приток раздражении из внешнего мира поступает в кору через аппараты ствола мозга.
Вторым важным источником поддержания постоянного тонуса коры являются импульсы, приходящие в кору от внутренних органов. Информация о состоянии организма (например, об уровне сахара в крови, являющемся показателем состояния голода), поступает в кору от аппаратов ствола мозга и гипоталамуса.
В работе «энергетического» блока мозга, кроме структур ствола мозга и гипоталамуса, участвуют аппараты древней («лимбической») коры, расположенные во внутренних (медиальных) отделах больших полушарий.
Нервные структуры первого блока связаны с корой с помощью волокон ретикулярной формации. Ретикулярная формация имеет как восходящие, так и нисходящие волокна. Посредством первых («восходящая активизирующая ретикулярная формация») осуществляется возбуждение коры импульсами, приходящими из верхних отделов ствола мозга. Посредством вторых («нисходящая активизирующая ретикулярная формация») осуществляются те влияния, которые высшие отделы мозга, и в частности его кора, оказывают на нижележащие отделы мозгового ствола. Например, с помощью «нисходящей ретикулярной формации» осуществляется передача в нижележащие отделы мозгового ствола программ поведения, которые возникают в коре.
Аппарат ретикулярной формации обеспечивает также третий источник поддержания тонуса коры, который связан со сложными замыслами и потребностями, формирующимися у человека при участии речи.
Ретикулярная формация имеет ряд особенностей строения и функционирования, благодаря которым обеспечиваются ее основные функции: во-первых, она состоит из восходящей и нисходящей частей. По волокнам восходящей ретикулярной формации возбуждение направляется вверх, оканчиваясь в расположенных выше образованиях (гипоталамусе, древней коре и новой коре). Волокна нисходящей ретикулярной формации имеют обратное направление: начинаясь от новой коры, они передают возбуждение к структурам среднего мозга и ствола мозга. Кроме того, нейроны ретикулярной формации работают по принципу «постепенного накопления возбуждения», т. е. возбуждение распространяется не отдельными импульсами, а градуально, постепенно меняя свой уровень и таким образом модулируя состояние всего нервного аппарата. И, наконец, к ретикулярной формации сходятся волокна (колатерали) от всех анализаторных систем, а также волокна из коры головного мозга и мозжечка. Наличие многочисленных связей в самой ретикулярной формации, конвергенция всех нервных путей на большей части ее нейронов создают дополнительные возможности широкого и одновременного распространения волн возбуждения в первичные, вторичные и третичные зоны коры, а также другие структуры мозга [Лурия А. Р.Основы нейропсихологии. М., 1973].
На основании результатов исследований Г. Мэгуна и Д. Моруцци (Mougoun Н., Morruzzi D.), опубликованных в 1949 г., принято считать, что у человека верхние отделы ретикулярной формации ствола мозга имеют связи с корой больших полушарий и регулируют уровень сознания, внимания, двигательной и психической активности. Эта часть ретикулярной формации получила название: восходящая неспецифическая активирующая система.
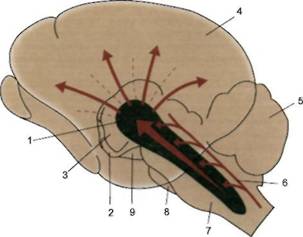 Ретикулярная формация ствола, ее активирующие структуры и восходящие пути к коре больших полушарий (схема).
Ретикулярная формация ствола, ее активирующие структуры и восходящие пути к коре больших полушарий (схема).
1 — ретикулярная формация ствола мозга и ее активирующие структуры; 2 — гипоталамус; 3 — таламус; 4 — кора большого мозга; 5 — мозжечок; 6 — афферентные пути и их коллатерали; 7 — продолговатый мозг; 8 — мост мозга; 9 — средний мозг.
К восходящей активирующей системе относятся ядра ретикулярной формации, расположенные, главным образом, на уровне среднего мозга, к которым подходят коллатерали от восходящих чувствительных систем. Возникающие в этих ядрах нервные импульсы по полисинаптическим проводящим путям, проходя через интраламинарные ядра таламуса, субталамические ядра к коре больших полушарий, оказывают на нее активирующее влияние. Восходящие влияния неспецифической активирующей ретикулярной системы имеют большое значение в регуляции тонуса коры больших полушарий, а также в регуляции процессов сна и бодрствования.
В случаях поражения активирующих структур ретикулярной формации, а также при нарушении ее связей с корой больших полушарий возникает снижение уровня сознания, активности психической деятельности, в частности когнитивных функций, двигательной активности. Возможны проявления оглушенности, обшей и речевой гипокинезии, акинетического мутизма, сопора, комы, вегетативного состояния.
В составе ретикулярной формации имеются отдельные территории, получившие в процессе эволюции элементы специализации — вазомоторный центр (депрессорные и прессорные его зоны), дыхательный центр (экспираторный и инспираторный), рвотный центр. Ретикулярная формация содержит структуры, влияющие на соматопсихо-вегетативную интеграцию. Ретикулярная формация обеспечивает поддержание витальных рефлекторных функций — дыхания и сердечно- сосудистой деятельности, принимает участие в формировании таких сложных двигательных актов, как кашель, чиханье, жевание, рвота, сочетанная работа речедвигательного аппарата, обшей двигательной активности.
Многообразны восходящие и нисходящие влияния ретикулярная формация на различные уровни нервной системы, которые «настраиваются» ею на выполнение той или иной конкретной функции. Обеспечивая поддержание определенного тонуса коры больших полушарий головного мозга, ретикулярная формация сама испытывает контролирующее влияние со стороны коры, получающей таким образом возможность регулировать активность собственной возбудимости, а также влиять на характер воздействий ретикулярной формации на другие структуры мозга.
Нисходящие влияния ретикулярной формации на спинной мозг сказываются прежде всего на состоянии мышечного тонуса и могут быть активирующими или понижающими мышечный тонус, что важно для формирования двигательных актов. Обычно активация или торможение восходящих и нисходящих влияний ретикулярной формации осуществляется параллельно. Так, во время сна, для которого характерно торможение восходящих активирующих влияний, происходит торможение и нисходящих неспецифических проекций, что проявляется, в частности, снижением тонуса мышц. Параллелизм влияний, распространяющихся от ретикулярной формации по восходящим и нисходящим системам, отмечается и при обусловленных различными эндогенными и экзогенными причинами коматозных состояниях, в происхождении которых ведущая роль отводится дисфункции неспецифических структур мозга.
При патологических состояниях взаимоотношение функций восходящих и нисходящих влияний может иметь и более сложный характер. При эпилептических пароксизмах, при горметоническом синдроме Давиденкова, возникающем обычно вследствие грубых поражений ствола мозга, угнетение функций коры больших полушарий сочетается с повышением мышечного тонуса.
Все это свидетельствует о сложности взаимоотношения функций различных структур ретикулярной формации, которые могут вести как к синхронным восходящим и нисходящим влияниям, так и к их нарушениям с противоположной направленностью. Вместе с тем ретикулярная формация является лишь частью глобальной интегративной системы, включающей лимбические и корковые структуры лимбико-ретикулярного комплекса, во взаимодействии с которыми и осуществляется организация жизнедеятельности и целенаправленного поведения.
Е. Д. Хомская подчеркивает, что функциональное значение первого блока в обеспечении психических функций заключается, во-первых, в регуляции процессов активации, в поддержании общего тонуса ЦНС, необходимого для любой психической деятельности (активирующая функция). Во-вторых, в передаче регулирующего влияния мозговой коры на нижележащие стволовые образования (модулирующая функция): За счет нисходящих волокон ретикулярной формации высшие отделы коры управляют работой нижележащих аппаратов, модулируя их работу и обеспечивая сложные формы сознательной деятельности [Актуальные проблемы нейропсихологии детского возраста / Под ред. Л. С. Цветковой. Москва — Воронеж, 2001.]
Нервная система всегда находится в состоянии определенной активности и для любого проявления жизнедеятельности обязательно ее наличие. Принято выделять несколько источников активности: в первую очередь, обменные процессы организма, лежащие в основе гомеостаза (белковый, углеводный и т. д.). Затем непосредственный приток информации, поступающей в организм из внешнего мира (от экстерорецепторов). Известно, что в состоянии сенсорной депривации человек впадает в сон, из которого его может вывести лишь поступление новой информации. Перечисленные источники активности свойственны и человеку, и животным. Но у человека помимо этого значительная часть активности обусловлена его планами, намерениями, программами. Формируясь в процессе сознательной жизни, они являются социальными по своему заказу и осуществляются при ближайшем участии сначала внешней, а потом внутренней речи[Лурия А. Р.Основы нейропсихологии. М., 1973].
Особенности патологических проявлений, связанных с нарушением функций ретикулярной формации, находятся в зависимости от характера, распространенности и выраженности патологического процесса и от того, какие именно отделы ретикулярной формации оказались в него вовлечены. Дисфункция лимбико-ретикулярного комплекса, и в частности РФ, может быть обусловлена многими вредными токсическими, инфекционными воздействиями, дегенеративными процессами в структурах мозга, расстройствами мозгового кровоснабжения, внутричерепной опухолью или травмой мозга.
При поражении ретикулярной формации снижается продуктивность всех ВПФ, в первую очередь — непроизвольного внимания и памяти, нарушается активность, сон. В случае массивных поражений стирается грань между сном и бодрствованием, человек находится в полусонном состоянии, у него страдает ориентировка во времени и месте. Отличительными диагностическими признаками поражения ретикулярной формации является одновременное снижение продуктивности абсолютно всех психических процессов, а также возможность частичной компенсации дефекта за счет усложнения задания. Вовлечение произвольных процессов и специальная мотивация позволяют ненадолго повысить эффективность психических процессов.
Таким образом, первый блок мозга участвует в обеспечении психической деятельности, в первую очередь в организации внимания, памяти, эмоционального состояния и сознания в целом. Кроме того, первый блок мозга участвует в регуляции эмоциональных (страх, боль, удовольствие, гнев) и мотивационных состояний. Лимбические структуры мозга, входящие в этот блок, занимают центральное место в организации эмоциональных и мотивационных состояний. В связи с этим первый блок мозга воспринимает и перерабатывает разнообразную интероцептивную информацию о состоянии внутренних органов и регулирует эти состояния.
Первый блок мозга, в состав которого входят аппараты верхнего ствола, ретикулярной формации и древней коры, обеспечивает общий тонус (бодрствование) коры и возможность длительное время сохранять следы возбуждения. Работа этого блока не связана с конкретными органами чувств и носит «модально-неспецифический» характер, обеспечивая общий тонус коры. Первый блок не участвует ни в приеме и переработке информации, ни в выработке программ поведения.
Второй блок — блок приема, переработки и хранения информации расположен в наружных отделах новой коры (неокортекса) и занимает ее задние отделы, включая в свой состав аппараты затылочной, височной и теменной коры. Структурно-анатомической особенностью этого блока мозга является шестислойное строение коры. Она включает первичные зоны (обеспечивающие прием и анализ поступающей извне информации), вторичные зоны (выполняющие функции синтеза информации от одного анализатора) и третичные зоны (основной задачей которых является комплексный синтез информации).
Отличительной особенностью аппаратов второго блока является модальная специфичность. Эксперименты по регистрации активности отдельных нейронов показали, что нервные клетки первичных зон отличаются высокой модальной специфичностью и узкой специализацией. Первое означает, что они реагирует на возбуждение только одной модальности (одного вида), например, только зрительное или только слуховое. Второе предполагает, что эти нейроны реагируют лишь на отдельный признак раздражителя одного вида (например, только на ширину линии или угол наклона и т. п.). Благодаря этому аппараты второго функционального блока мозга выполняют функции приема и анализа информации, поступающей от внешних рецепторов и синтеза этой информации.
Второй блок обеспечивает анализ и синтез сигналов, приносимых органами чувств из внешнего мира, т.е. связан с приемом, переработкой и хранением получаемой человеком информации. Он включает структуры, расположенные в задних отделах коры головного мозга: теменной, височной и затылочной области, и в отличие от первого блока, имеет модально-специфический характер.
Структуры этого блока представляют собой центральные (корковые) отделы анализаторов. Эти центральные структуры воспринимают зрительную, слуховую и тактильную информацию, перерабатывают или «кодируют» ее и сохраняют в памяти следы полученного опыта. Корковые отделы зрительного анализатора расположены в затылочной, слуховые — в височной, тактильно-кинестетические — в теменной области.
В этих отделах коры кончаются волокна, идущие от соответствующих воспринимающих, рецепторных зрительных, слуховых и тактильных аппаратов. Те зоны коры, куда непосредственно приходят волокна от периферических чувствующих органов, носят название первичных, или проекционных, зон. Зоны коры примыкающие к проекционным, называются вторичными, или проекционно-ассоциативными, зонами.
Механизм работы второго блока следующий. Основу работы блока составляет принцип иерархического построения каждой зоны коры головного мозга. Информация, поступающая от зрительного, слухового и кожного рецепторов в первичные зоны коры, дробится там на огромное число составляющих ее признаков. В проекционных зонах коры находятся высокоспециализированные нейроны, реагирующие только на определенные частные признаки раздражении. Например, в проекционной зоне затылочной зрительной коры существуют нейроны, которые реагируют только на движение светящейся точки от центра к периферии или от периферии к центру, либо только на плавные изогнутые линии и т. п. Такие же специализированные нейроны существуют и в височной слуховой и тактильной теменной коре. Они позволяют дробить возбуждение на отдельные элементы, делая его доступным для дальнейших операций.
Над каждой первичной зоной надстроены вторичные, или проекционно-ассоциативные, зоны коры. Волокна, поступающие сюда, не приходят непосредственно от периферического рецептора. Они либо несут обобщенные импульсы, либо приходят во вторичные зоны коры из первичных. Эти нейроны не отличается такой тончайшей специализацией, как нейроны первичных зон. Они реагируют не на отдельные дробные признаки, а чаще всего на комплекс модально-специфических: зрительных, слуховых, тактильных раздражителей. Некоторые нейроны имеют мультимодальный характер, реагируя на раздражения различных модальностей. Значение этих вторичных зон состоит в том, чтобы объединять и перекодировать информацию, приходящую от нижележащих подкорковых ядер или от первичных зон коры.
Первичные зоны чувствительной коры осуществляют функцию раздробления, т.е. анализа поступающей информации на ее составные части, в то время как вторичные зоны этих же отделов коры несут функцию объединения – синтеза или сложной переработки доходящей до субъекта информации.
Первичные и вторичные зоны коры не исчерпывают корковых аппаратов анализируемого блока. Над ними надстроены аппараты третичных зон коры, которые также называют зонами перекрытия корковых концов отдельных анализаторов, которые обеспечивают комплексную работу этого блока. Третичные зоны коры головного мозга являются специфически человеческими образованиями.
Третичные зоны коры созревают очень поздно в онтогенезе. Их основная функция заключается в объединении информации, приходящей в кору от различных анализаторов. Больные с поражением этих отделов мозговой коры воспринимают раздельно и последовательно зрительные и слуховые раздражители и не могут объединить их в одновременные схемы. Эти больные испытывают затруднения при ориентировке в пространстве, путают направления, не могут различать правую и левую стороны, имеют трудности в понимании сложных грамматических структур и в логических операциях. Все это свидетельствует, что третичные зоны коры являются аппаратом, необходимым для наиболее сложных форм переработки и кодирования получаемой информации.
А. Р. Лурия выделяет основные законы построения коры, входящей в состав второго блока мозга [Лурия А.Р.Основы нейропсихологии. М., 1973]. Давайте познакомимся с ними подробнее.
1) Закон иерархического строения корковых зон. Согласно этому закону соотношение первичных, вторичных и третичных зон коры осуществляет все более сложный синтез информации. Более сложно организованные зоны коры обеспечивают более сложные функции. А. Р. Лурия подчеркивает, что соотношение первичных, вторичных и третичных зон у взрослого и у ребенка различно. Для нормального развития вторичных зон у ребенка необходимо, чтобы были сформированы первичные, а для развития третичных — вторичные зоны. Поэтому поражение первичных зон в раннем детстве может приводить к грубым нарушениям в развитии вторичных и, тем более, третичных зон. У взрослого же человека, при сформированных зонах коры, третичные, наиболее организованные, управляют функцией ниже лежащих вторичных и первичных зон. Поэтому у взрослого человека взаимодействие зон коры осуществляется сверху вниз. В данном случае поражение первичных зон не приводит к заметным нарушениям психических функций и может компенсироваться работой расположенных рядом структур.
2) Закон убывающей модальной специфичности иерархически построенных корковых зон предполагает, что по мере перехода от первичных зон к третичным снижается проявление их модальной специфичности. Первичные зоны каждой из долей мозга, входящих во второй блок мозга, обладают максимальной модальной специфичностью (благодаря громадному числу нейронов с высокодифференцированной, модально-специфической функцией). Вторичные зоны, в которых преобладают верхние слои с ассоциативными нейронами, обладают модальной специфичностью в значительно меньшей степени. Еще меньше модальная специфичность характерна для третичных зон описываемого блока («зоны перекрытия» корковых концов различных анализаторов). Таким образом, этот закон описывает переход от дробного отражения частных, модально-специфических признаков к синтетическому отражению более общих схем воспринимаемого мира.
3) Закон прогрессивной латерализации функций объясняет связь функций с определенным полушарием (по мере перехода от первичных зон к третичным зонам). Первичные зоны обоих полушарий мозга равноценны. На уровне вторичных зон часть функций, выполняемых левым и правым полушариями, остаются одинаковыми, но часть функций левого полушария уже отличаются от функций, выполняемых правым полушарием мозга. Функции же третичных зон левого полушария уже коренным образом отличаются от функций аналогичных зон правого полушария мозга.
Е. Д. Хомская указывает, что при поражении аппаратов второго блока мозга нарушение функций зависит от того, какие именно зоны пострадали. При поражении первичных зон возникает нарушение восприятия отдельных признаков воспринимаемого раздражителя одной модальности (слепое пятно, гемеанопсия, нарушение тон-шкалы, анестезия и т. д.). При поражении вторичных зон коры наблюдается нарушение синтеза отдельных признаков воспринимаемого раздражителя в целостный образ одной модальности (агнозии, афазии). Поражение третичных зон приводит к нарушению комплексного синтеза раздражений, поступающих от разных анализаторов, что проявляется в нарушении ориентировки в пространстве. Причем, согласно закону прогрессивной латерализации, при поражении третичных зон правого полушария нарушается предметная ориентировка в пространстве, а при поражении аналогичных зон левого полушария — страдает символическая ориентировка в пространстве. [Хомская Е. Д. Нейропсихология. М., 1987.]
Второй функциональный блок мозга составляют анализаторы, или сенсорные системы. Сенсорные системы или анализаторы, по И.П.Павлову, – это часть нервной системы, состоящая из воспринимающих элементов – рецепторов, получающих стимулы из внешней или внутренней среды, нервных путей, передающих информацию от рецепторов в мозг, и тех частей мозга, которые перерабатывают эту информацию. Рецепторы – концевые образования афферентных нервных волокон, воспринимающие раздражения из внешней (экстерорецепторы) среды или внутренней (интерорецепторы) среды организма и преобразующие энергию раздражителей в возбуждение, передаваемое в анализаторные зоны коры головного мозга.
Все известные части сенсорных систем включают как минимум следующие компоненты: 1) детекторы стимула – специализированные рецепторные нейроны; 2) первичный воспринимающий центр, куда сходится информация от группы детекторных блоков; 3) один или большее число вторичных воспринимающих и интегрирующих центров, получающих информацию от первичных воспринимающих центров. Взаимодействие этих центров и создает “восприятие”.
Каждый анализатор настроен на определенную модальность сигнала и обеспечивает описание всей совокупности признаков воспринимаемых раздражителей.
Все анализаторы тесно связаны, в связи с этим в коре большого мозга осуществляется анализ и синтез, выработка ответных реакций, регулирующих любые виды деятельности человека. Анализатор – это многоуровневая система с иерархическим принципом ее конструкции. Основанием анализатора служит рецепторная поверхность, а вершиной – проекционные зоны коры. Каждый уровень этой конструкции представляет собой совокупность клеток, аксоны которых идут на следующий уровень, исключение составляет верхний уровень, аксоны которого выходят за пределы данного анализатора. Взаимоотношения между последовательными уровнями анализаторов построены по принципу “дивергенции-конвергенции”. Дивергенция – способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. Конвергенция – схождение различных импульсных потоков от нескольких нервных клеток к одному и тому же нейрону. Чем выше нейронный уровень анализаторной системы, тем большее число нейронов он включает.
Проекционные зоны анализаторных систем занимают наружную поверхность новой коры задних отделов мозга. Сюда входят зрительная затылочная, слуховая височная и соматосенсорная теменная области коры. В корковый отдел этого функционального блока включаются также первичные структуры вкусовой, обонятельной, висцеральной чувствительности.
Первичная зона коры – область мозга, к которой подходят чувствительные волокна, начинающиеся в периферических органах чувств. Значительная часть этих нейронов первичных зон коры обладает высочайшей функциональной специфичностью. Так нейроны зрительных областей избирательно реагируют на определенные признаки зрительных раздражителей: одни – на оттенки цвета, другие – на направление движения и т.д. Таким образом, при восприятии стимула не происходит равномерного возбуждения всей проекционной зоны: активируются, главным образом, те нейроны, к которым поступает информация от возбужденных периферических элементов – рецепторов. Этот первичный этап нередко обозначают как сенсорный анализ.
Принципиально меняется характер обработки информации при переходе к вторичным ассоциативным зонам коры. Являясь модально специфическими, эти зоны, по образному утверждению А.Р.Лурии, превращают топическую проекцию раздражения в функциональную организацию.
Вторичные проекционные зоны коры располагаются вокруг первичных зон, как бы надстраиваясь над ними. Они осуществляют синтез раздражителей, функциональное объединение различных анализаторных зон, принимая непосредственное участие в обеспечении видов познавательной деятельности. Для этих нейронов характерно детектирование сложных признаков раздражителей, однако, при этом сохраняется модальная специфичность, соответствующая нейронам первичных зон. Поэтому предполагается, что усложнение детекторных селективных свойств нейронов вторичных зон может происходить путем конвергенции на них нейронов первичных зон.
Первичные и вторичные зоны, согласно И.П.Павлову, составляют центральную часть, или ядро анализатора в коре, нейроны которого характеризуются избирательной настройкой на определенный набор параметров раздражителя и обеспечивают механизмы тонкого анализа и дифференцировки раздражителей. Взаимодействие первичных и вторичных зон обусловливает согласованное содружество процессов возбуждения и торможения, что создает основу для динамического межанализаторного взаимодействия, осуществляемого в ассоциативных зонах коры.
Ассоциативные области (третичные зоны) коры осуществляют сложные надмодальные виды психической деятельности – символической речевой, интеллектуальной. Ассоциативные зоны расположены на границе затылочной, височной и заднетеменной коры (заднеассоциативная область) и во фронтальных отделах коры (переднеассоциативная область).
В высших сенсорных (вторичных и ассоциативных) зонах коры действует закон убывающей специфичности, который является обратной стороной принципа иерархической организации нейронов-детекторов в специфической подкорке и проекционных зонах коры. В нем отражается переход от дробного анализа частных модально-специфических признаков раздражителя к синтезу более общих “схем” воспринимаемого. Закономерным является то, что, несмотря на убывающую специфичность высших сенсорных полей коры, они являются в функциональном отношении более совершенными образованиями. Они выполняют функцию интеграции сложных комплексных раздражителей, им свойственна пластичность, они подвержены “неспецифической” активации со стороны модулирующих систем.
Третий функциональный блок мозга — блок программирования, регуляции и контроля сложных форм деятельности. Он связан с организацией целенаправленной, сознательной психической активности, которая включает в свою структуру цель, мотив, программу действий по достижению цели, выбор средств, контроль за выполнением действий, коррекцию полученного результата. Обеспечению этих задач и служит третий блок мозга.
Аппараты третьего функционального блока мозга расположены кпереди от центральной лобной извилины и включают в свой состав моторные, премоторные и префронтальные отделы коры лобных долей мозга. Лобные доли отличаются очень сложным строением и большим числом двусторонних связей со многими корковыми и подкорковыми структурами. Отличительной особенностью этого блока является проведение процессов возбуждения от третичных зонах к вторичным, затем к первичным; отсутствие модально-специфических зон (состоит из аппаратов только двигательного типа); наличие обширных двусторонних связей не только с нижележащими образованиями ствола мозга, но и со всеми остальными отделами коры больших полушарий.
Сознательная деятельность человека только начинается с получения и переработки информации, а кончается она формированием намерений, выработкой соответствующей программы действий и выполнением этих программ во внешних (двигательных) или внутренних (умственных) актах. Аппарат создающий намерения, вырабатывающий программы действий, осуществляющий эти программы и контроль за протекающими действиями, сличая эффект выполняемого действия с исходными намерениями работает на базе передних отделов мозга и их лобными долями, составляющими третий блок головного мозга.
Как и задние отделы мозга, передние имеют теснейшие связи с нижележащими образованиями ретикулярной формации. Здесь представлены как восходящие, так и нисходящие волокна ретикулярной формации. Нисходящие волокна проводят импульсы, сформированные в лобных долях коры, в нижележащие структуры, тем самым регулируя активность организма в соответствии со сформированными в коре намерениями.
Передние отделы мозга также построены по иерархичному принципу и включают первичные, вторичные и третичные зоны. Первичной, или проекционной, зоной передних отделов мозга является передняя центральная извилина - моторная область коры; над ней надстроено вторичное, премоторное поле; еще выше расположены образования собственно лобной или префронтальной области коры - поля 9, 10, 11, 46 Бродмана.
Долгое время значение лобных отделов коры в организации поведения не поддавалось научному определению. Это связано с тем, что поражение лобных долей мозга не приводит к нарушениям элементарных движений, не вызывает ни параличей, ни расстройств чувствительности, ни нарушений речи. Поэтому лобные доли коры расценивали как «немую зону», не имеющую каких-либо специальных функций. Эти взгляды изменились, когда исследователи стали подходить к мозгу как к сложнейшей саморегулирующей системе, которая создает сложные программы поведения, регулирует протекание двигательных актов и осуществляет контроль над ними.
Лобные доли мозга, обладающие мощными связями с восходящей и нисходящей ретикулярной формацией, оказались аппаратом, обладающим мощной активизирующей ролью. Во время сложной интеллектуальной работы, требующей повышенного тонуса коры, в лобных долях возникает повышенное число синхронно возбуждающихся совместно работающих нейронных ансамблей. Эти синхронно работающие структуры сохраняются во время сложной интеллектуальной работы и исчезают после ее прекращения. Лобные отделы мозга поддерживают тонус коры, необходимый для выполнения поставленной поведенческой задачи.
Двустороннее поражение лобных долей мозга приводит к тому, что больные оказываются не в состоянии прочно удерживать намерения, сохранять сложные программы действий, тормозить не соответствующие программам импульсы и регулировать деятельность. Они не могут устойчиво концентрировать свое внимание на поставленной перед ними задаче и легко отвлекаются от ее выполнения.
Другой важной функцией лобных долей мозга является осуществление постоянного контроля над протекающей деятельностью. Больные с поражением лобных долей мозга не могут сличить результаты своих действий с исходным намерением, лишены возможности осознавать собственные ошибки и исправлять их. Лобные доли мозга обеспечивают механизм саморегуляции деятельности человека.
По своей структуре и функциональной организации моторная кора относится к первичным, премоторная — к вторичным, а префронтальная — к третичным зонам коры больших полушарий. Поэтому они выполняют функции, характерные для этих зон. Нейроны моторной коры передают возбуждение к мышцам, отсюда начинается большой пирамидный путь. Именно эти зоны имеют ярко выраженную соматотопическую организацию, что наглядно проиллюстрировал Пенфилд («двигательный человечек»). Премоторная кора обеспечивает двигательные программы, т. е. объединяет отдельные движения в единую кинетическую мелодию. Префронтальные отделы играют решающую роль в формировании намерений, программ, в регуляции и контроле наиболее сложных форм поведения человека. Они состоят из мелкозернистых клеток с короткими аксонами и обладают мощными пучками восходящих и нисходящих связей с ретикулярной формацией. Поэтому могут выполнять ассоциативную функцию, получая импульсы от первого блока мозга и оказывать интенсивное модулирующее влияние на образования ретикулярной формации, приводя ее активирующие импульсы в соответствие с динамическими схемами поведения, которые формируются непосредственно в префронтальной (лобной) коре. Префронтальные отделы фактически надстроены над всеми отделами мозговой коры, выполняя функцию общей регуляции поведения [Лурия А. Р.Основы нейропсихологии. М., 1973].
При поражении третьего функционального блока мозга характер нарушения функций связан с тем, какой именно аппарат поврежден. При поражении моторной коры затрудняется проведение возбуждения к конкретным мышцам (наблюдаются парезы и параличи отдельных групп мышц). Поражение премоторной коры приводит к нарушению синтеза отдельных движений в единое целое (распад двигательных навыков), поражение префронтальных отделов проявляется в нарушении сознательной целенаправленной деятельности. Исследования А. Р. Лурия показали, что в этом случае целенаправленное поведение заменяется «полевым» поведением, нарушается программа выполняемой деятельности, лобный больной не может подобрать адекватные средства деятельности, отсутствует контроль за ее выполнением и коррекция ошибок. При этом наблюдается персеверация (повторяемость) и стереотипизация движений.
Можно резюмировать, что первым функциональным блоком коры головного мозга является энергетический блок головного мозга, он играет важную роль в регуляции состояний активности коры и уровня бодрствования. Этот блок построен по типу неспецифической нервной сети, которая осуществляет свою функцию путем постепенного, градуального изменения состояний и не имеет непосредственного отношения ни к приему и переработке поступающей извне информации, ни к выработке намерений, планов и программ поведения. Этот функциональный блок, расположенный в основном в пределах ствола мозга, медиобазальных отделах, существенно отличается от аппаратов третьего и второго функционального блока мозга.
Второй функциональный блок приема, переработки и хранения сенсорной информации расположен в задних отделах полушарий и включает в свой состав зрительные затылочные, слуховые височные и общечувствительные теменные отделы коры головного мозга и соответствующие подкорковые структуры.
Аппараты этого, как и следующего блока имеют иерархическое строение, распадаясь на первичные проекционные зоны, которые принимают информацию и дробят ее на мельчайшие составные части, вторичные проекционно-ассоциативные зоны, которые обеспечивают кодирование, т.е. синтез этих составных частей и превращают соматотопическую проекцию в функциональную организацию, и третичные зоны или зоны перекрытия, обеспечивающие совместную работу различных анализаторов и выработку надмодальных символических схем, лежащих в основе комплексных форм познавательной деятельности.
Указанные иерархически построенные зоны коры разбираемого блока работают по принципам убывающей модальной специфичности и возрастающей функциональной латерализации. Оба эти принципа и обеспечивают возможность наиболее сложных форм работы мозга, лежащих в основе наиболее высоких видов познавательной деятельности человека, связанных с трудом и с участием речи в организации психических процессов.
Третий функциональный блок мозга - блок программирования, регуляции и контроля за протекающей деятельностью. Аппараты третьего функционального блока мозга расположены в передних отделах больших полушарий, спереди от центральной извилины. Он состоит целиком из аппаратов эфферентного двигательного типа, однако сам находится под постоянным притоком информации из аппаратов афферентного сенсорного блока. Следующая важнейшая черта, отличающая работу третьего функционального блока от афферентного, состоит в том, что процессы здесь идут в нисходящем направлении, начинаясь с наиболее высоких - третичных и вторичных зон коры. Здесь в высших отделах интегративно-пускового блока формируются двигательные программы, а затем переходят к аппаратам низших моторных образований - первичным корковым зонам; стволовым и спинальным двигательным ядрам.

