




По оценкам разных специалистов, современные моховидные представлены 20-25 тыс. видов, распространенных на всех континентах. Особенно это относится к Северному полушарию, где на больших площадях (на болотах, иногда в лесах) моховидные доминируют в растительном покрове. Эти растения предпочитают наиболее увлажненные места, однако произрастают и на каменистых склонах гор, в пустынях, на сухих открытых скалах, в тундрах, но не встречаются в морях, на сильно засоленных почвах, в ледниках и сыпучих песках.
Подавляющее большинство моховидных - низкорослые многолетние травянистые растения размером от 1 мм до нескольких сантиметров, реже до 60 см и более. У большинства из них тело расчленено на стебель и листья, но встречаются и слоевищные формы.
Листостебельные представители моховидных имеют ряд признаков, которые отличают их от остальных высших растений. У них отсутствуют корни; прикрепление к субстрату (частично и всасывание воды) осуществляется одним или многочисленными ризоидами - выростами эпидермы. Поглощение воды происходит главным образом нижней частью стебля или телом всего растения (сфагновые мхи).
Вегетативные органы сформированы ассимиляционной, проводящей, запасающей и покровной тканями. Однако в отличие от других высших растений у моховидных отсутствуют настоящие сосуды и механические ткани. На стебле зеленых мхов по спирали расположены бесчерешковые листья, имеющие цельную пластинку с жилкой или без нее.
Это единственная группа растений, эволюция которой связана с регрессивным развитием спорофита и преобладанием в жизненном цикле гаметофита. Спорофит моховидных – спорогон - состоит из коробочки и ножки, нижняя расширенная часть которой - стопа - внедряется в ткань гаметофита и доставляет спорофиту воду и питательные вещества. Спорофит, таким образом, лишен самостоятельности и полностью зависит от гаметофита.
Основная функция спорофита - образование спор, которые в благоприятных условиях прорастают. С момента прорастания споры начинается развитие гаметофита - полового поколения моховидных. Прежде всего развивается разветвленная нитчатая (у большинства мхов) или пластинчатая (у сфагнума) протонема, на которой закладываются почки. У одних мхов из почек протонемы формируются пластинчатые слоевища, у других - листостебельные побеги гаметофита (взрослого мха), на которых развиваются органы полового размножения - антеридии и архегонии.
Размножаются мхи спорами, могут размножаться и вегетативным путем - отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса: Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
2 Класс печеночные и антроценовые мхи.
Класс Печеночные мхи ( Hepaticopsida ) насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha) (рис 95).

1– таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6 – вертикальный разрез через женскую подставку (а – архегоний)
Рисунок 95 - Цикл воспроизведения маршанции
Гаметофит имеет темно-зеленое слоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляется ризоидами. На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках. Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии. На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Антоцеротовые мхи, или антоцеротопсиды (Anthocerotae, Anthocerotopsida), класс моховидных. Слоевище розетковидное, диаметром 1-3 см, реже лентовидное, с волнистыми или изрезанными краями, тёмно-зелёное, плотно прилегающее к почве. Многочисленные, слегка изогнутые щетинковидные зелёные спорогоны выcотой 2-3 см придают антроценовым мхам своеобразный вид. При созревании спорогоны чернеют, растрескиваются по двум скручивающимся створкам, выбрасывая споры, и постепенно вытягиваются (у некорых видов – до 10 см и более) благодаря меристеме в основании. В клетках таллома и спорогонов содержится один или несколько хлоропластов с пиреноидами – характерная особенность этих мхов. Одно семейство, 4-5 родов, свыше 300 видов, преимущественно в тропиках и субтропиках. К роду антоцерос (Anthoceros) относится около 200 видов, в основном – тропических.
3 Класс листостебельные мхи
Класс Листостебельные мхи (Bryopsida, или Musci).Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи. Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichiumcommune), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии (рис. 96).
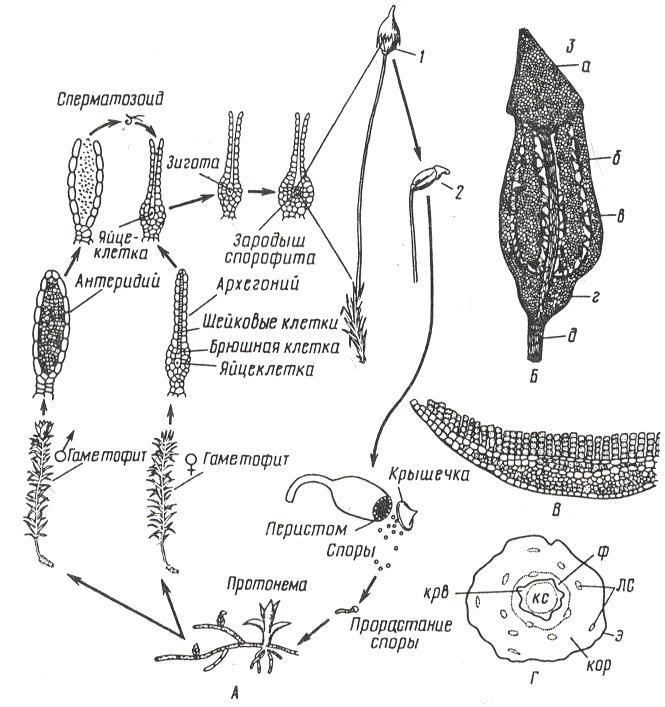
А – цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы)
Рисунок 96 – Цикл развития кукушкина льна
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора - небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой - гаплоидное поколение - гаметофит. Коробочка на ножке - диплоидное поколение - спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae). К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum).
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток:
1) живых ассимилирующих, длинных и узких, хлорофиллоносных;
2) гиалиновых – мертвых, лишенных протопласта.
Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел псилофиты.
Псилофиты – это чрезвычайно своеобразная группа примитивных современных высших споровых растений. Представлена двумя родами – Псилот (Psilotum) и тмезиптерис (Tmesipteris), объединенными в одно семейство Писилотовые (Psilotaceae) порядка Псилотовидные (Psilotales) класса Псилотоподобные (Psilotopsida) (рис. 97).

Рисунок 97 – Представитель псилофитов
Род Псилот включает 2 вида, мезиптерис – 10, обитающих во влажных тропических и частично субтропических областях обоих полушарий. Они играют очень скромную роль в сложении современного растительного покрова. Это многолетние травянистые растения, наземные или эпифитные. Произрастают на перегнойной почве, в трещинах скал, у основания стволов деревьев.
Эпифиты поселяются на стволах древовидных папоротников, саговников, пальм.
Для псилотовых характерен комплекс чрезвычайно примитивных признаков, которые позволяют проследить направления эволюции растительного мира.
Псилотовые лишены корней. Их подземные органы, достигающие длины 1 м и более, дихотомически ветвятся, морфологически соответствуют ризомоидам силурийских риниофитов. В отличие от корневищ, ризомоиды лишены каких-либо чешуй (видоизмененных листьев), а покрыты многочисленными ризоидами (на корневищах развиваются корневые волоски). Эпидермальные клетки ризомоида и ризоиды покрыты кутикулой. В клетках коры содержатся гифы эндотрофных грибов, проникающие через ризоиды. Благодаря микоризным грибам псилотовые ведут частично сапрофитный образ жизни. Отсутствие корней – главное своеобразие псилотовых. Корни отсутствуют во все периоды их жизни (даже на стадиях развития зародыша). Отсутствие корня у них не является результатом редукции, как это свойственно другим растениям в результате приспособления к особым условиям обитания. У псилота и тмезиптериса отсутствие корней – это первичный признак, сближающий с риниофитами. С риниофитами их сближает и ряд других примитивных черт анатомического строения. В частности, проводящая система ризомоида гаплостелическая.
Наземные оси достигают высоты 10–100 см у псилота и 5–40 см у тмезиптериса; у эпифитных особей обычно свисающие, у наземных – часто прямостоячие. Ветвление дихотомическое. Более обильное у псилотов, причем самого примитивного равнодихотомического типа. Многие тмезиптерисы не ветвятся. Проводящая система трехгранных наземных осей псилота представлена актиностелью, у тмезиптериса возникает более или менее ясно выраженная сифоностела. Прото- и метаксилема образованы трахеидами различного морфологического строения (спиральные, кольчатые, лестничные). В метаксилеме псилота встречаются даже точечно-поровые трахеиды. Ксилему окружает флоэма, снаружи стель окружена эндодермой.
К периферии от стели располагается кора с листовыми следами, внешние слои которой представлены фотосинтезирующей тканью, состоящей из тонкостенных паренхимных клеток. Эпидерма, покрывающая наземные оси с устьицами, имеет утолщенные наружные тангентальные стенки, снабженные кутикулой. Таким образом, основным органом ассимиляции у псилотовых является стебель.
Примитивным признаком псилотовых является отсутствие типичных листьев. В верхней части стеблей псилотов имеются мелкие чешуевидные придатки, беспорядочно расположенные. Они лишены устьиц и проводящих пучков (хотя в коре стебля имеются листовые следы). Чешуевидные придатки псилотов напоминают энационные листья астероксилона (класс Плауноподобные), у которого проводящий пучок лишь подходил к основанию листа. Более листовидный характер имеют эти придатки у тмезиптерисов. В нижней части стебля они также мелкие и чешуевидные как у псилотов. Лишены проводящего пучка. Выше по стеблю они крупнее, имеют вид ланцетных, низбегающих по стеблю зеленых пластинок, заканчивающихся пленчатым отростком. Они снабжены одиночным проводящим пучком, имеют устьица с обеих сторон. Чешуевидные придатки псилотов в своем развитии остановились на уровне девонских форм, а у тмезиптерисов – достигли развития современных плауновидных.
Спороносные структуры псилотовых также примитивного строения, с чертами некоторой специализации. Спорангии образуют синангии (трехспорангиальные у псилотов и двуспорангиальные у тмезиптерисов), (рис. 8). Образование синангиев являеся признаком специализации. Однако эта черта эволюционной продвинутости невысокого ранга. Тенденция к образованию синангиев наблюдалась уже среди ринифотов – ярравия. Синангии псилотов располагаются терминально, на вершинах коротких веточек, имеющих вид толстых ножек; у тмезиптерисов – на листоподобных придатках. Свойственное псилотовым терминальное расположение синангиев является примитивным, и у других современных растений не встречается.
Спорангии также примитивного строения, толстостенные (стенка состоит из 4–6 слоев клеток). Значительная часть спорогенной ткани разрушается и превращается в питательную массу для спороцитов (материнских клеток спор) и спор.
Псилотовые являются морфологически равноспоровыми растениями. Споры многочисленные почковидные, с прозрачными оболочками и имеют сетчатую поверхность. Зрелые спорангии вскрываются продольной щелью.
Как и спорофиты, не менее своеобразны гаметофиты (заростки) псилотовых. Они также характеризуются рядом примитивных особенностей. Они цилиндрические, на концах слабо неравно дихотомически разветвленные, наземные или подземные, размером 1,5–2 см. Они лишены хлорофилла, сопрофитные и очень похожи на фрагмент ризомоида спорофита. Питаются микоризно за счет гиф симбиотических грибов. От заростка отходят многочисленные одноклеточные ризоиды. Своеобразной особенностью заростков псилотовых является наличие слабо развитой примитивной стели. По мнению некоторых исследователей она представляет результат редукции когда-то хорошо развитых гаметофитов предков писилотовидных, имевших стелярное строение. Заростки обоеполые. Антеридии и архегонии рассеяны по всей поверхности. Антеридии поверхностные, не погружены в ткань заростка. В них развиваются спиральные многожгутиковые сперматозоиды. Архегонии брюшком погружены в ткань заростка, с короткой выступающей шейкой.
Формирующийся после оплодотворения зародыш, как и у всех высших споровых растений, развивает гаусторию, абсорбирующую питательные вещества из тканей заростка. Остальная часть зародыша еще на стадии недифференцированных тканей дихотомически делится. За счет одного апекса развивается надземная ось, а из другого – ризомоид или вторая надземная ось. Таким образом, у псилотовых еще нет генетически закрепленной дифференциации органов. На зародышевой стадии у них не образуется ни корня, ни первичного листа, ни подвеска. Молодой спорофит долго связан с внешне похожим на него заростком.
Псилотовые могут размножаться вегетативным путем. Ответвления ризомоидов, выходя на поверхность, магут развивать новые надземные оси.
Как отмечено, они не играют заметной роли в сложении современного растительного покрова. Невелико их практическое значение в жизни человека. Некоторые формы псилотов культивируются как оригинальные декоративные растения. Благодаря равно дихотомическому ветвлению надземная часть растения приобретает правильный контур. Темно-зеленые стебли украшены ярко-желтыми синангиями. Они представляют научный интерес как резко обособленная, длительно эволюционирующая группа примитивных современных растений.
Филогенетическое происхождение псилотовых и их место в системе растительного мира не совсем ясное. Во многих современных системах они рассматриваются как первично примитивыне формы, как прямые потомки риниофитов. С последними их сближает наличие ризомоида, отсутствие корней, дихитомическое ветвление, относительно примитивное внутреннее и внешнее строение надземных осей и спороносных структур. Существует также мнение о них как о формах вторично упрощенных. Поскольку палеонтологический материал отсутствует, проблема остается не решенной. По последним данным прослеживается тесное филогенетическое родство с наиболее примитивными современными папоротниками – ужовниковыми.
Основная литература:
1 Еленевский А.Г., Соловьев М.П., Тихомиров В.Н. Ботаника: систематика высших, или наземных, растений. 2 изд. - М.: Academіa, 2001. - 429 с.
2 Нестерова С.Г. Лабораторный практикум по систематике растений. - Алматы: Қазақ ун-ті, 2011. - 220 с.
3 Родман А.С. Ботаника. – М.: Колос, 2001. - 328 с.
Дополнительная литература:
1 Абдрахманов О.А. Систематика низших растений. – Караганда: Изд-во КарГУ, 2009. - 188 с.
2 Билич Г.Л., Крыжановский В.А. Биология. Т. 2: Ботаника. - М.: Оникс 21 век, 2002. - 543 с.
3 Ишмуратова М.Ю. Систематика и интродукция растений (курс лекций). - Караганда: РИО Болашак-Баспа, 2015. - 100 с.
4 Тусупбекова Г.Т. Основы естествознания. Ч. 1. Ботаника. – Астана: Фолиант, 2013. – 321 с.
Контрольные вопросы:
1 Какое поколение преобладает у моховидных?
2 В чем заключаются отличие между мхами и псилофитами?
3 Какая группа среди споровых растений является наиболее примитивной?
4 Где развивается спорофит у моховидных?
5 В чем заключаются примитивные признаки развития и жизненного цикла у псилофитов?
6 Каким образом могут размножаться мхи и псилофиты?
Лекция 18. Отдел Плауновидные (Lycopodiophyta), Хвощевидные (Equisetophyta)
План лекции:
1 Отдел Плауновидные.
2 Отдел Хвощевидные.

