




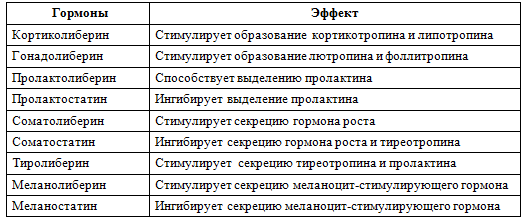
Адам ағзасының реттеуші жүйелерінің ішінде гипоталамо-гипофизарлы жүйе ең жоғары сатыда түр. Гормондардың гипоталамус пен гипофизда түзілуі жэне бөлінуі бір-бірімен тығыз байланысты. Мысалы, гиполаламустың клеткаларында ерекше нейропептидтер-либериндер (рилизинг-факторлар) синтезделеді. Мидың белгілі бір орталықтарының қозуына жауап ретінде гипофизде аяқталатын гипоталамустың жүйке клеткаларының аксондарынан либериндер бөлінеді де, гипофиздың клеткаларынан сэйкес тропты гормондардың: соматолибериннің, тиролибериннің, кортиколибериннің, фоллилибериннің, люлибериннің, пролактолибериннің, меланолиберинің синтезін жэне бөлінуін ынталандырады.
Либериндермен қатар гипоталамуста нейропептидтердің басқа бір тобы - статиндер де түзіледі, олар гипофизарлы гормондардың: соматостатинның, пролактостатинның, меланостатинның жэне т.б. синтезі мен бөлінуін ингибиторлайды. Либериндер мен статиндердің түзілуі гипоталамустың тубероинфундибулярлы бөлігінде іске асырылады. Аркуатты ядроларда-лю жэне фоллилибериндер, сүр төмпешіктің нейрондарындакортиколиберин, орталық төмпешік аймаңында-тиролиберин, вентромедиальды ядроның үстінде пролактолиберин жэне пролактостатин синтезделеді.
Тиреолиберин – пироглутамин қышқылынан, гистидиннен, пролинамидтен тұратын триптид; пептидті байланыспен байланысқан. N және C соңды аминқышқылдарында бос NH 2, COOH топтары жоқ.
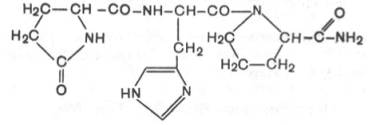
Люлиберин – тізбектелген 10 аминқышқылынан тұратын декапептид. С-соңды аминқышқыл глицинамид.
Соматостатин – 14 аминқышқыл қалдығынан тұратын тетрадекапептид. Құрамында цистеин қалдықтарының арасында дисульфидті байланыс бар.
Соматолиберин – табиғи көздерден алынбаған, тек декапетидтің химиялық синтезі жүргізілген. Ол өсу гормонының секрециясы мен синтезіне стимулирлеуші әсер етеді.
Меланолиберин – химиялық құрамы окситоциннің ашық тізбегіне аналогті:
Н - Цис-Тир-Иле-Глн-Асн-Цис-ОН
Меланостатин - трипептид Пиро-Глу-Лей-Гли- NH 2, немесе пентапептид түрінде болады. Меланолиберин меланотропиннің синтезі мен секрециясына стимулирлеші, меланостатин ингибирлеуші әсер етеді.
3.АКТГ, СТГ гормондары химиялық табиғаты, биологиялық әсері.Адренокортикотропты гормон (АКТГ) немесе кортикотропин аденогипофиздың базофиль-ды клеткаларында синтезделеді жэне оның бөлінуі гипоталамустың кортиколиберинімен ын-таландырылады. Химиялық табигаты бойынша АКТГ-39 аминқышқылды қалдықтан түратын пептид болып табылады. 1-24 қалдықтарының N-соңғы фрагменті барлық омыртқалыларда бірдей болатыны анықталған. АКТГ-ның С-соңгы тізбегі, эсіресе 25-33 бөлігі эртүрлі жану-арларда эртүрлі болады. Дэл осы фрагмент АКТГ-ның бөтен препараттарын беруге жауап ретінде ағзаның қажетсіз реакцияларының дамуына жауапты. Қазіргі кезде клиникада АКТГ-ның 1 -24 фрагменті болып табылатын синактен деп аталатын синтетикалық препараты пайдаланылады. Табиғи АКТГ-ның сандық жэне сапалық гормональды белсенділігін сақтай отыра, синактенді адам мен жануарларға бергенде иммундық жауапты тудырмайды.
АКТГ ыдыраған кезде жаңа гормональды эсері бар 2 пептид түзілуі мүмкін екені
анықталған:
1-13 фрагментінде альфа-МСГ қасиеті бар,
18-39 фрагменті-инсулинің бөлінуін ынталандырушы. Кортикотропты гормон бүйрек үсті бездерінің кьізметін бақылайды, онда холестериннен кортикостероидты гормондардың, эсіресе глюкокортикоидтың түзілуін ынталандырады, сонымен қатар андрогендердің жэне аз молшерде альдостеронның синтезіне эсер етеді.
АКТГ-ның кортикостероидтердің биосинтезіне эсер ету механизмін Гаррен жэне оныц қызметкерлері (1971) анықтаған. АКТГ бүйрек үсті бездері қабатының клеткаларының ре-цепторларымен байланысады жэне аденилатциклазды-протеинкиназды механизм арқылы ри-босомада ерекше белоктың синтезін іске қосады. Бүл белок холестеринның Р450 цитохром-мен өзгешелік байланысуын жэне оның липидті тамшылардан митохондрияларға жеткізілуін қамтамасыз етеді. Осымен бір мезетте, циклды АМФ-пен белсендірілген протеинкиназа холестерин эфирлерін ыдыратушы эстеразаны белсендіреді, "а" фосфорилазаны белсендіреді жэне пентозды циклда глюкозо-6-фосфаттың тотығуын ынталандыру арқылы НАДФН2-ні жинақтайды. Стероидты гормондардың биосинтезінің өзіне тэн ерекшелігі - митохондриялар мен микросомаларда бірқатар кезектілікпен жүретін прегненолонның түзілуі арқылы холестериннің бүйірлі тізбегінің қысқару процестері жэне стероидтар молекулаларының гидроксильдену реакцияларының жүруі болып табылады. Бул процестер гидролазалар немесе аралас типті оксидазаларга жататын ерекше ферменттік жүйелермен іске асырылады жэне НАДН2 - ден оттегіге электрондарды тасымалдаумен тіркесе жүретін ферменттердің айрықша тізбегін қалыптастырады. Қорыта келгенде, синтезделетін стероидты гормондарга қосылатын гидроксильды топқа оттегі атомының енуі жүреді:
НАДФ2 —> ФАД—> Адренодоксин —> Р450 —> 02
1 І
холестерин—> прегненолон
І
кортикостерон
Селье жэне басқа да авторлардың көптеген зерттеулері эртүрлі стресс кезінде жалпы адаптациялық синдромның іске асуының маңызды үйымдастырушыларының бірі-гипоталамо-гипорфизарлы-бүйрек үсті безді жүйесі болып табылатынын көрсетті:
-Ағзаға кез-келген стрессорлы эсер кезінде гипоталамустың-кортиколиберинды, гипофиздың-АКТГ-ны жэне бүйрек үсті бездерінің-глюкокортикоидтарды коп молшерде бөлуі байқалды. Гипоталамусты-гипофизарлы-адреналды жүйенің белсендірілуі дабыл кезеңінде өтеді жэне белгілі бір интервалдарда стрессорлы фактордың күшіне тура пропор-ционалды болады.
Соматотропин немесе гипофиздың сомототропты гормоны (СТГ) немесе осу гормоны-полипептидтік тізбегі 191 аминқышқылдық қалдықтан түратын белок. Бүл түр өзгешелік қасиеті бар бірден-бір белок болып табылады. Өсу гормонының бөлінуі гипоталамустың сомотолиберині мен соматостатинінің бақылауында болады, өз кезегінде олардың бөлінуі қандағы глюкозаныц деңгейіне тэуелді. Соматостатин СТГ-ның бөлінуін тежейді жэне үйқы безінің инсулин мен глюкагонды бөлуін шектейді. Соматотропинның көптеген биологияльщ эсері бар, ол сүйектің, ішкі мүшелердің салмагының өсуін, үйқы безінің глюкагон мен ин-сулинды бөлуін ынталандыру арқылы көрінеді. Алайда, эсер ету аумағының кең болуына карамастан, осу гормонының бауырдыц клеткаларында рецепторлары бар, онда ол соматомединдердің түзілуін ынталандырады - бүл белоктық қосылыстар арқылы самото-тропты гормон барлық тканьдерге реттеуші эсер етеді.
Соматомединдердің 7 түрі бар: Al, А2, Bl, В2, ВЗ, В4, С. А жэне С типті соматоме-диндер шеміршек клеткаларының бөлінуін, ДНҚ, РНҚ, белок, коллаген синтезін жэне протеогликандардыц сульфирленуін күшейтеді. С соматомедины май жэне бүлшьщ ет тканьдеріне эсер етеді, бүл жерде олардьщ глюкозаны сіңіруін ынталандырады жэне май тканінде липолизды тежейді, бауырда глюконеогенезді белсендіреді. В соматомединдері жүйке тканінің клеткаларында ДНҚ жэне белок синтезін ынталандырады. Соматотропты гормон фибробластардың пролиферациясын күшейтеді жэне олардьщ проколлаген мен хондроитин-сульфаттарды синтездеуін жеделдетеді. Әсіресе түтікті сүйектердің эпифизарлы бөлігінде орналасқан шеміршек тканьдері соматотропинге сезімтал болады. Ересек ағзада СТГ-ның өсу әсері азаяды да, анаболитикалық эсері түгелімен сақталады. Соматотропты гормон эпидермис, жүйке жэне т.б. осу факторы тэрізді өсудің ткань- өзгешелік жергілікті факторларының синтезінынталандырады. Сонымен қатар л имфоидтыклеткалардыңантиденелердісинтездеуін ынталандырады жэне қабыну реакциясының жоғары қарқындылыгына эсер етеді. Жалпы, соматотропты гормон ағзаның өзгеріп түратын қажеттілігіне байланысты, алмасу процестерін реттеп, бағыттап отырады жэне заттар алмасуына мынандай эсер етеді: 1.Өсу гормонының белокты алмасуга эсері.
- аминқышқылдарды пайдалануды жэне белокты синтезді жеделдету арқылы жүмсақ тканьдердің өсуін жэне ішкі мүшелердің көлемін арттырады.
-ДНҚ синтезін жэне иіеміршек тканьдерінің пролиферациясын ынталандырады, -бауырда белок синтезін күшейтеді жэне қан плазмасында белоктар деңгейін арттырады -лимфоидты клеткаларда антиденелердің синтезін ынталандырады, 2.Липидтер алмасуына эсері 2 кезеңнен тұрады:
-алғашқы 3-6 сағатта (бірінші кезең) осу гормоны липолиз бен май қышқылдарының тотығуын тежейді, бауырда липидтер синтезін ынталандырады,
-8-12 сағаттан кейін қанға СТГ өткеннен соң (екінші кезең) май депосынан липидтер мобилизациясы жүреді, қанда НЭЖК жэне үшацилгицеридтердің деңгейі артады, май қышқылдарының бета-тотығуы жэне кетон денелерінің синтез, жеделдейді.
3.Соматотропты гормонның көмірсулар алмасуына эсері:
-гипергликемия мен гликогеннің мобилизациясын жеделдетуге экелетін глюкагон мен серотонинның бөлінуін ынталандырады
-гексокиназаның белсенділігін тежеу арқылы глюкозаның тотығуын төмендетеді, -бауырда глюконеогенезді ынталандырады,
-қанға инсулинның бөлінуін күшейтеді жэне осы гормонды ыдыратушы фермент-инсулиназаны белсендіреді. Осы бойынша СТГ Лангерганс аралшаларының d-клеткаларының гиперплазиясына жэне дегенерациясына эсер етеді, яғни диабетогендік эсер көрсетеді.
4.Өсу гормонының су-түзды алмасуға эсері:
-қанда бейорганикалық фосфаттардың деңгейін арттырады,
-зэрмен Са^шығарылуын арттырады,
-тканьдер мен қанда Mg++ Na+, К+ иондарының деңгейінің сақталуын қамтамасыз етеді.
Соматотропты гормон жэне онымен функциональды байланысқан рилизинг-факторлар жэне соматомединдер стрессорлы гормондар тобына жатады. Ағзадағы эртүрлі стрессорлық эсер кезінде олардың қан плазмасындағы деңгейі артады. Өсу гормонының жас өсіп келе жатқан ағзада жеткіліксіз молшерде бөлінуінде өсудің тежелуі байқалады, ол бойдың өспей қалуына экеледі (гипофизарлы нанизм). Гипофизарлы нанизммен ауыратын науқастарда дене бітімінің дүрыс пропорциялары сақталады жэне ақыл-естің дамуы өзгермейді. Мұндай бойдың өспей қалуы ергежейлілерде кездеседі. Жас ағзада өсу гормонының шектен тыс бөлінуі гигантизмге экеледі. Ал егер өсу гормонының гиперпродукциясы ағзаның өсуі тоқтаған кезде болса, онда ол акромегалия деп аталатын жагдайды тудырады. Акромегалия қаңканың, бас сүйектің, иектің, қолдың, аяқтың, буындардың фиброзды тканьдерінің жэне кейбір жүмсақ тканьдердің (тілдің) пропорциональды емес өсуі арқылы көрініс береді. Ерекше көрінетін акромегалия қантты диабеттің симптомдарымен қатар жүреді. Әйел адамдарда жүктіліктің соңғы айларында плацентаның соматотропиндерді бөлуіне байланысты, элсіз физиологиялық өтпелі акромегалия байкалады.
4, Нейрогипофиз гормондары – химиялық табиғаты және биологиялық әсері. Гипофиздың артқы бөлігінің гормондары- вазопрессин мен окситоцин гипоталамустың супраоптикалық жэне паравентрикулярлы ядроларында синтезделеді, ол жерден нейросе-креторлы жолмен гипофиздың арткы бөлігіне түсіп, сол жерде жинақталды және нейроген-ды қоздыргыштар эсер еткен сайын қанға бөлінеді. Сол себепті вазопрессин мен окситоцин нейрогипофизарлы гормондар деп аталады. Вазопрессин мен окситоцин химиялық табигаты бойынша бір-бірінен 2 аминқышқылы арқылы ерекшеленетін, 9 аминқышқылды қалдықтан түратын циклды нонапептид болып табылады.

Вазопрессин немесе антидиуретикалық гормон (АДГ)-тканьдық сүйықтықтың осмостық қысымының артуына "жауап" ретінде гипоталамустың нейросекреторлы гранулаларынан бөлінеді жэне нейрогипофизбен секрецияланады. Вазопрессинның басты эсер ететін орны-бүйрек нефроны, оның дистальды тармақталған түтіктері жэне жинақтау түтікшелері жэне жинақтау түтігі болып табылады. Бүл жерде аденилатциклазды-протеинкиназды механизм арқылы натрий иондарының, хлоридтердің жэне бастапқы зэрден судың қайтадан қанға реабсорбциясының белсенді тасымалын қамтамасыз ететін мембраналық белоктардың белсендірілуі жүреді (антидиуретикалық эсер). Вазопрессин эпителиальды клеткалардың өткізгіштігіне эсерін 3' -5' -АМФ-тың кемегімен жүзеге асырады. Вазопрессин клеткалық мембрананың апикальды қабығының "қуысын " үлғайтады да, су үшін мукозды кедергіні азайтады, оның клетка ішіне диффузды ағынын күшейтеді. Су клеткадан серозды кабат арқылы қанға өтеді. Жоғары концентрацияда аденилатциклазаны ингибиторлайтын Са++ иондары вазопрессинның гидроосмостық эсерін тежейді. Сонымен бірге, вазопрессин вазо-прессорлы эсер көрсетеді, ол перифериялык артериолалар мен жүрек артериолаларының тегіс мускулатурасының жиырылуын тудырады. Несеп қуығы мен ішектің тегіс мускулатурасының жиырылу белсенділігін арттырады.
Бүйректе судың қайта сіңірілу деңгейі өте жоғары жэне антидиуретикалық гормонның қызметімен қамтамасыз етіледі. Вазопрессинның жеткіліксіз бөлінуі кезінде (қантты емес диабетте) түтікшелердің дистальды бөліктерінде жэне нефронның жинақтау түтігіндегі судың реабсорбциясы күрт төмендейді. Бүл кезде тэуліктік диурез артады. Кей жағдайда тэуліктік диурез 30 литрға тең болады (қалыпты жағдайда 1,5-2,0 литр). Бүл қанның осмостық қысымының жоғарылауына, қан қысымының төмендеуіне, клетка тургорының азаюына, терінің құрғауына экеледі.
Қанның осмостық қысымының маңызды реттеушісі ретінде вазопрессинді бөлуші гипоталамо-нейрогипофизарлы комплекс осмостық процестердегі кез-келген өзгерістер мен соған байланысты гемодинамика мен диурез өзгерістеріне сезімтал болады. Бүл процестер барысындағы кез-келген ауытқулар теріс кері байланыс механизмы бойынша вазопрессиннің деңгейінің өзгеруіне экеледі. Мысалы: қан кеткен кездегі қан плазмасының көлемінің бар болтаны 8%-ға азаюы-вазопрессинның секрециясының тез арада артуына экеледі. Белокты ерітінділерді жіберу, шөлдеу, тұзды, глюкозаны мөлшерден тыс қабылдау, қан қысымының төмендеуі, гипоксия жэне т.б. вазопрессиннің секрециясына ынталандырушы эсер көрсетеді. Бүл жагдайларда вазопрессинның көп мөлшерде бөлінуі плазмада су мөлшерінің артуына, қанның осмостық қысымының қалпына келуіне, циркуляциядағы сүйықтықтың мөлшерінің жэне қан қысымы мөлшерінің артуына себепкер болады. Керісінше, диурездың тежелуі, тұзы аз диета, қан қысымының жоғарылауы, гипероксия антидиуретикалық гормонның бөлінуін тежейді.
Вазопрессин мөлшерден тыс бөлінгенде ағзада судың жинақталуы байқалады, қан қысымы жоғарылайды, жұмсақ ісіктер, бастың ауруы, олигоурия пайда болады. Зәрдің салыстырмалы салмагы артады. Бүл өзгерістер Пархон ауруы (Пархон синдромы) кезінде байқалады.
Жатырдың, сүт безінің, шэует шылбыры жолдарының тегіс мускулатурасының клеткаларында окситоцинның рецепторлары бар. Бүл гормон жатырдың жиырылуының күшті ынта-ландырушысы ретінде босану процесінің инициациясы мен дүрыс өтуінде ең маңызды орын алады.
Лактацияланатын сүт безінен сүттің эвакуациясында окситоцин ерекше орын алады. Бүл гормон емізу актісіне рефлекторлы түрде нейрогипофизбен бөлінеді жэне сүт безінен сүттің шыгуына септігін тигізеді де, ерекше миоэпителиальды клеткалардың жиырылуын тудырады. Бездік клеткалардан сүттің коп молшерде бөлінуін қамтамасыз ете отыра, окситоцин жанама түрде лактация процесін ынталандырады. Окситоцин босану кезеңінде жеткіліксіз бөлінсе, жатырдың жиырылу белсенділігі элсіз болуы мүмкін, ол тек ана емес, бала үшін де өте қауіпті жагдай болып табылады. Окситоцин мөлшерден тыс болганда ауыр салдары бар жедел босану орын алуы мүмкін. Жүктілік кезінде окситоцинның жогары деңгейі түсікке экелуі мүмкін.
5. гипофиздің гонадотропты гормондарының әсер ету механизмі. Гонадотропты гормондар тобына -ФСГ (фолликулин ынталандырушы гормон) жэне лю-теонизде (ЛГ) уші гормон жатады.
Химиялық табиғаты бойынша ЛГ жэне ФСГ - альфа жэне d-типті 2 суббірліктерден тұратын күрделі белоктар - гликопротеидтер болып табылады. Бүл гормондардың биологиялық эсері бета-суббірлікпен байланысты. Осы гормондардың бөлінуі гипоталамустың рилизинг-факторларының: люлибериндер, фоллибериндердің бақылауында болады. Гонадотропин-дер мен олардың бөлінуін бақылайтын лю-фоллитропиндер адамның дамуының эмбрио-нальды кезеңінен бастап түзіле бастайды. Алайда гонадалар қызметінің қалыптасуындағы жэне жыныстық дамудағы олардың мэні бүл кезенде шектеулі болады. Постэмбрионалды кезеңде, эсіресе препубертантты кезенде андрогендер немесе экстрогендердің инициация-лаушы эсеріне байланысты олардың қызметі күшейеді. Аталық бездің жэне аналық бездің гонадотропиндердің эсеріне сезімталдығы артады.
Ересек жануарлар мен адамда гипофиздың гонадотропиндерді бөлуін реттеуші гипоталамустық бөлімдер 2 орталықтан түрады: өтпелі мидың преоптикалық зонасын-да орналасқан циклды орталықтан жэне аркуатты зонада орналасқан ациклды орталықтан. Люлиберинды бөлуші ациклды орталықтың клеткалары ерлерде жэне үрғашыларда жүмыс істейді де, гонадотропиндердің секрециясының базальды деңгейін анықтайды. Циклды орталық тек үрғашыларда жүмыс істейді де, жүйке импульсациясы есебінен аркуатты зона клеткаларында рилизинг-фактордың бөлінуінің эр мезеттегі (периодтық) өзгерістерін анықтайды да, сол арқылы эйелдердің жыныстық циклдарының іске асуын қамтамасыз етеді. Гипоталамустың циклды қызметі тек жыныстық жетілу аяқталғаннан соң ғана жүзеге аса бастайды. Гипоталамустық орталықтардың аналық немесе аталық типі бойынша дифферен-цировкасы эмбриогенез кезінде жыныс гормондарымен немесе постэмбриональды дамудың ерте кезеңдерінде (неонатальды кезеңде) детерминделеді.
Лютеонизирлеуші гормонның (лютропин) құрамында 88 аминқышқылды қалдықтан түратын а-суббірлік жэне 119 аминқышқылды қалдықтан түратын бета-суббірлік бар. Жыныстық жетілмеген үрғашыларда ЛГ фолликулалардың өсуі мен жетілуін жэне олар-да эстрогендер мен прогестиндердің биосинтезін ынталандырады. ЛГ-овуляцияның негізгі индукторы. Оның овуляцияға эсері аналық бездерде гонадотропиндердің эсерінен түзілетін простагландиндер арқылы жүзеге асады. Простагландиндердің қатысуымен ЛГ фолликулада транскрипция жэне трансляция деңгейінде протеолитикалық ферменттердің синтезін күшейтеді, ол фолликулярлы қабаттың бүзылуы мен аналық безден жүмыртқа клеткасының қүрсақ қуысына шығуына экеледі. ЛГ-ның циклды бөлінуі мен осы кезде ФСГ мен пролактинның коп молшерде бөлінуі эсерінен жетілген фолликул клеткаларының оған жогары сезімталдыгы овуляцияны тудырады, ол ЛГ-ның бөлінуінен кейін 8-50 сағаттан соң пайда болады. Қанға ЛГ-ның бөлінуінің максимумы эйелдерде овариальды циклдың 14-17 күнінде-овуляция кезеңінде байкалады. ЛГ-сары дене қалыптасуының жэне олардың проге-стеронды синтездеуінің басты индукторы болып табылады. Лютеинизация процесіне ЛГ-ның эсері пролактинмен пермисссияланган. Осыдан пролактинның екінші -лютеотропты гормон (ЛТГ) атауы пайда болды. ЛТГ-ЛГ рецепторларының түзілуін күшейтеді жэне сол арқылы ЛГ-ның томен концентрациясына лютеинды клеткалардың жогары сезімталдығын қамтамасыз етеді. Аталған гормондардың, эсіресе ЛГ-ның эсерінен пайда болган сары дененің лютеинды клеткалары негізінен прогестиндерді (прогестеронды) синтездейді де, гипоталамус-гипофиз-гонадалар жүйесінің жэне циклдың лютеин кезеңінде жыныс жолының функциональды жалпы жағдайын анықтайды. Сары денелер бөлетін прогестиндер сүтқоректілерде жатырды жүктілікке дайындайды, гипоталамустың циклды орталығының белсенділігін жэне аналық безде фолликулдың жетілуін тежейді. Егер жүктілік болмаса, сары денелер инволюциялана-ды да, оларда прогестерон синтезі тежеледі. Ерлер агзасында ЛГ-тестостерон синтезі бойынша Лейдиг клеткаларының өсуі мен пролиферациясын ынталандырады. ЛГ-онтогенездің постнатальды кезеңінде андрогендер биосинтезінің негізгі реттеушісі. ЛГ- перифериялық тканьдердің андрогендер мен экстрогендерге сезімталдыгын арттырады. Сонымен, лютропин (ЛГ)-овуляцияның жэне аналық бездерде сары дененің түзілуінің, аталық бездерде ин-терстициальды клеткалардың өсуінің негізгі ынталандырушысы, сонымен қатар экстраген-дер, прогестиндер жэне гонадальды андрогендер биосинтезінің ынталандырушысы болып табылады.
Фоллитропин-фолликулин ынталандырушы гормон (ФСГ), ЛГ сияқты гликопротеидты табигаты бар 2 суббірліктерден түрады. ФСГ-ның бета-суббірлігінде ЛГ молекуласының осы типтес суббірлігіне Караганда көмірсулық қалдықтар 4 есе коп болады.
ФСГ-үрғашыларда овариальды фолликулдардың өсуін жеделдетеді, оларды ЛГ эсеріне сенсибилизациялайды. Жыныс гормондарымен катар ФСГ ерлерде Лейдиг клеткаларының ЛГ-ның эсеріне сезімталдығын арттырады, сперматогенездің кейбір кезеңдерін жэне тести-кулдарда Сертоли клеткаларының өсуін ынталандыруга қабілетті.
ЛГ жэне ФСГ жануарлардың жыныстық дамуына экстрагональды эсер етеді. Атап айтқанда, гонадотропиндер перифериялық тканьдерді жыныс стероидтарының эсеріне сенсибилизациялайды, ерлердің жыныс сферасының мүшелерінде, бауырда жэне бүйректе андрогендер метаболизмінің ферменттерінің синтезіне эсер етеді. Гонадотропиндердің овуляция мен фолликулдардың жетілуін ынталандыратын қабілеті кеңінен қолданыс табуда. Аналық бездердің поликистозымен ауыратын эйелдерді емдеу үшін гонадотропиндерді пайдаланады (овуляция болмай, репродуктивті қызметі бүзылғанда). Ауыл шаруашылығында гонадотроп-ты гормондардың препараттары қойды көп төлдету үшін, үрық шашуды ынталандыру үшін балық шаруашылығында жэне қүстарда жүмыртқа салуды жақсарту үшін пайдаланылады.
Пролактин-лактогенді гормон немесе гипофиздың лютеотропты гормоны (ЛТГ) мономерлі белок болып табылады, ол қүрамы жағынан СТГ-ға үқсас, 199 аминқышқылды қалдықтардан кұралган. Пролактин молекуласында 3 дисульфидтік байланыстар бар. ЛТГ бөлінуі гипоталамустың пролактолиберині мен пролактостатиннің бақылауында болады. СТГ сияқты, пролактин де өсуді реттеуге қатысады. Омыртқалылардың төменгі класында бүл гормон СТГ эсерін қайталайды. Сүтқоректілер мен қүстарда пролактин ішкі мүшелерде анаболитикалық процестерді ынталандырады, шаш пен қанаттың өсуін, май бездерінің өсуі мен олардың белсенділігін күшейтеді, эритропоэзды ынталандырады. Адам мен сүтқоректілерде пролактинның сүт бездерінің өсуі, дифференцировкасы жэне қызметін реттеуде маңызы зор. Жоғары омыртқалыларда, соның ішінде сүтқоректілерде пролактинның жалпы даму процесіне эсері жете зерттелмеген. Пролактин өсу процестерін ынталандырады жэне ағзаның дамуын жэне көптеген түрлі клеткалар дифференци-ровкасын ингибиторлайды деп есептеледі. Сонымен бірге, пролактин-сүт бездерінің дифференцировкасының жэне гонадалардың гонадотропиндер эсеріне сезімталдығының дифференцировкасының маңызды гормональды факторларының бірі болып табылады. СТГ, тиреоидты гормондар, эстрогендер жэне баска да гормондармен катар пролактин сүтқоректілердің үрғашыларында сүт бездерінің біртіндеп гормонға тэуелді дифферен-цировкасын қамтамасыз етеді. Жүктілік кезінде сүт бездері дамуының қорытынды кезеңі өтеді-сүт бездерінің альвеолярлы аппаратының белсенді түрі қалыптасады жэне өсуі ын-таланады, ол сүттің өзгешелік факторын синтездеуге қабілетті қүрамын қалыптастырады жэне лактацияның алғашкы кезеңдерінде молозиво (коллострум), сонан соң сүттің өзін бөледі. Сүт бездерінің лактацияның алдындағы кезеңіндегі дифференцировкасының басты инициаторлары бастапқы кезде прогестерон мен хорионды соматомаммотропин (ХСТ), сонан соң пролактин (ЛТГ) болып табылады. Сүттің өзгешелік факторларының синтезі мен лактацияның өзі жүктілік кезінде басталады. Бүл кезде аз молшерде көбінесе молозиво бөлінеді. Сүттің көп бөлінуі босанган соң алғашқы күндерден-ақ басталады жэне сүт бөлінуінің ынталандырушысы пролактин болып табылады. Пролактин-босанғаннан кейін сүттің бөлінуінің басты ынталандырушысы. Босанганнан кейінгі кезеңде гипофиздың про-лактинды бөлуінің ынталандырылуы эстрогендердің жоғары деңгейімен жэне босанған эйелдің агзасындагы прогестиндердің төменгі деңгейімен анықталады. Бүл гормондардың ЛТГ секрециясына эсері гипоталамус жэне гипофиз арқылы іске асырылады. Сүт безінің альвеолярлы клеткаларында пролактин аденилатциклазды механизм арқылы протеинкиназаны белсендіреді, ол өз кезегінде сүт белоктарының синтезі (казеино-ген, бета-лактальбумин) жэне УДФ-галактоза мен глюкозадан лактозаның синтезін қамтамасыз етуші ферменттері галактозил-трансфераза ("А белогы") жэне альфа-лактальбулин ("В белогы") синтезі бойынша транскрипция мен трансляцияның ядролық процестерін іске қосады. Пролактинның бұл эсерлері жүктілік кезінде сүт факторларының биосинтезі мен бездік клеткалар диффе-ренцировкасын анықтайтын гормондармен күшейеді немесе бағытталады, атап айтқанда СТГ күшейеді жэне глюкокортикоидтармен пермиссияланады. Глюкокортиидтардың көп бөлінуінің себебі-эйел ағзасында пайда болатын босану кезіндегі жэне босанганнан кейінгі стресс жағдайы. Сонымен қатар прогестерон, эсіресе тестостерон сүттің өзгешелік компоненттерінің түзілуін тежейді. Пролактин лютеинизация процесіне ЛГ-ның эсерін пермиссиялайды (бағыттайды). Осыдан пролактинның екінші атауы-лютеотропты гормон (ЛТГ) пайда болды. ЛТГ-ЛГ рецепторларының түзілуін күшейтеді де, сол арқылы лютеинды клеткалардың ЛГ-ның томен концентрациясына жогары сезімталдығын қамтамасыз етеді. Сонымен қатар пролактин жүмыртка басу, балапандарды эртүрлі жеммен қоректендіру мен оларды үйрету инстинктының көрінуін қамтамасыз етеді. ЛТГ-қүстарда ғана емес, сонымен қатар сүт қоректілер мен басқа да омыртқалыларда аналық инстинктың, үрпағын күту инстинктың қалыптасуының маңызды ынталандырушысы (стимуляторы) болып табылады. Гормонның бүл эсерлері жыныстық дамумен жэне көбеюмен таңдаулы байланысты. Пролактин-қалқанша безінің гормондарының антагонисты. Амфибияларда пролактинның физиологиялық мэні-олардың личинка кезеңінде дене көлемінің өсуін жэне метаморфоза процестерінің тежелуін қамтамасыз ету болып табылады. Ал тиреоидты гормондардың қызметі-личинкалар денесінің жалпы көлемінің өсуін тежеу жэне метаморфозаны ынталандыру болып табылады. Сонымен, пролактинның биологиялық әсерінің аумағы өте кең, эйелдер мен ерлер ағзасында өзіндік ерекшеліктері бар. Әйел ағзасында пролактин сүт бездерінің дамуын жэне лактация процесін ынталандырады, эктодермальды құрылымдар мен ішкі мүшелердің өсуін, сары дененің түзілу процесін ынталандырады жэне оның функ-циональды белсенділігін қамтамасыз етіп, аналық инстинктың көрінуіне қажетті шарттар-ды тудырады. Ерлер ағзасында эктодермальды қүрылымдар мен ішкі мүшелердің өсуін ынталандырады, екіншілік жыныс белгілерінің қалыптасуына жыныс гормондарының эсерін күшейтіп, антигонадотропты әсер көрсетеді.
6. Қалқанша бездің йодтирониндері (тироксин, трииодтиронин) құрылысы, түзілуі, әсер ету механизмі, жасушаның зат алмасуына және биоэнергетикалық процестерге әсері.
Қалқанша безінің гормондарына иодтирониндер - тироксин (Т4) жэне трииодтиронин (ТЗ) жатады, олар тирозин аминкышқылының туындылары болып табылады. Олар Жер бетіндегі ең коне ағзаның бірі - көк-жасыл балдырларда табылған.

Иодтирониндердің биосинтезі қалқанша безінің фолликулаларында жүреді. Биосинтетикалық процестерге фолликула қабатын түзуші эпителиалды клеткалар ғана емес, сонымен қатар қуысты толтыру-шы коллоид та қатысады. Қалқанша безінің иодтирониндерінің синтезінің көпкомпонентті процесі мынадай кезеңдерден түрады:
- қан иодидтерінің калқанша безінде фиксациясы жэне элементарлы иодқа дейін тотығуы.
- өзгешелік белок-тиреоглобулинның синтезі жэне оның тирозилды қалдықтарының элементарлы иодпен иодталуы
- тиреоглобулин молекуласында иодталған тирозилды қалдықтардан иодтирониндердің түзілуі.
- тиреоглобулиннен тиреоидты гормондардың (Т4,ТЗ) бөлінуі жэне олардың қанға
өтуі.
Бүл процестердің барлығы гипоталамо-гипофизарлы жүйенің бақылауында болады. Атап айтқанда, гипофиздың тиреотропины (ТТГ) аденилатциклазды механизм арқылы тиреоглобулин синтезін іске қосады, қаннан қалқанша безіне иодидтердің белсенді тасымалдануын жэне енуін күшейтеді, тиреоглобулин молекуласында тирозинды радикалдардың иодталуын ынталандырады, қалқанша безі коллоидына тиреоглобулинның пиноцито-зын ынталандырады, коллоидтың сұйылу процесін белсендіреді, фолликулярлы клеткалар лизосомаларына тиреоглобулин жэне коллоидты тамшылардың қосып алынуын, тиреоглобулинның протеолизын жэне қанға иодтирониндердің бөлінуін белсендіреді.
Адреналин жэне Е2 простагландины қалқанша безінде цАМФ концентрациясын арттырады. Олар тиреотропин сияқты эсерлер көрсетеді. Қалқанша безі фолликулаларының иодты қосып алу процесі бір-біріне тэуелсіз 2 кезеңнен тұрады:
- каннан иод иондарының қалқанша безі эпителиалды клеткаларына энергияға тэуелді белсенді тасымалдану кезеңі, ол тотыгу фосфорильдену жэне тканьдік тыныс алу процестерімен байланысты (йод иондарының белсенді тасымалдануы қалқанша безінің клеткалық мембраналарының цАМФ-тәуелді фосфорильдену жолымен жылдамдайды);
- йодидты иондардың йодидпероксидазамен молекулярлы йодқа дейін тотығу кезеңі:
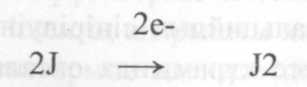
Қалқанша безінде йодтың жинақталу процесіне йод- пероксидазды реакцияны тежеуші қосылыстар кедергі жасайды. Оларға құрамында күкірт бар, тиреостатикалық зат-тар: метил-тиоурацил, пропилтиоурацил, 5-винил-2-тиооксазолидон, тиомочевина жатады. Тиреоглобулин биосинтезі белгілі бір ретпен өтетін 4 кезеңнен тұрады:
- рибосомаларда (полисомаларда) полипептидті тізбектердің түзілуі;
- полипептидті тізбектердің гликозильденуі,
- цитозольда тиреоглобулинның төртіншілік құрылымының қалыптасуы,
- тиреоглобулинның тирозилды қалдықтарының иодталуы жэне қалқанша безі фолликулдарының коллоидында иодталған йодтирониндердің түзілуі.
Тиреоидты гормондардың биосинтезінің негізгі кезеңі - тиреоглобулинның иодтиро-зилды қалдықтарының қалыптасуы жэне моноиодтирозин мен дийодтирозиннің түзілуі, йодтирозинның екі молекуласының одан ары конденсациясы мен тиреоглобулинның құрамында тироксинның (Т4) жэне трииодтиронинның (ТЗ) түзілуі.
Иодтиронинның түзілу процесінде иодтирозиндер молекуласынан дегидроаланин бөлінеді:

Тиреоидты гормондар биосинтезінің соңғы кезеңі-тиреоглобулиннен тироксин мен трийодтиронинның бөлінуі. Бул процесс пиноцитоздан басталады-иодталған тиреогло-булины бар фолликулярлы коллоид тамшыларын эпителиальды клеткалардың қосып алуы жэне олардың одан ары лизосомалармен қосылуы (фаголизосомалардың түзілуі). Фаголизосомдарда тироксин мен трийодтиронинның бөлінуімен тиреоглобулин протеоли-зы өтуі, бұдан соң тиреоидты гормондар қанға бөлінеді. Бөлінетін Т4 жэне ТЗ қатынасы 4:1-гетең,яғни олардың биосинтезінің деңгейіне сэйкес. Қанда тиреоидты гормондар қан плазмасының өзгешелік тасымалдаушы белоктарымен: тироксин байланыстырушы глобу-линмен, I, II преальбуминдермен байланысады жэне эффекторлы клеткаларға жеткізіледі.
Белоктармен байланысуына орай тиреоидты гормондар бірнеше күн ішінде қанда өзгермеген күйінде болады.
Тироксинның дейодталуы тиреоидты гормондардың мүше-нысаналар деңгейінде биологиялық эсерінің көрінуінде маңызды орында түр. Тироксин молекуласынан 5 орындагы (Т4) йод атомы бөлінген кезде 3',3,5-трииодтиронинның (ТЗ) түзілуі жүреді, оның тироксинге қарағанда белсенділігі оте зор. Тироксин дейодталуының басқа да өнімдері: 3\5\5-трийодтиронин, 3,3\5'-ТЗ жэне 5\3,5-ТЗ-биологиялық белсенді емес, сол себепті ТЗ реверсивті түрлері деп аталады.
Иодтирониндер үшін мүше - нысаналар - бауыр, жүрек, бүйрек, қаңқа бұлшық еттері болып табылады. Т4 жэне ТЗ -ке май жэне жүйке тканьдерінің сезімталдығы оте томен. Эффекторлы мүшелердің клетка-нысаналарында тиреоидты гормондар тканьдердің өсуін, дифференцировкасын жэне энергетикалык алмасуды реттейді жэне бүл жагдайда алмасу процестеріне екі жақты эсер етеді:
- біріншіден, цитозольды рецепторлар арқылы клетканың генетикалық аппаратына эсер етіп, транскрипцияны жэне жүзден артық ферменттердің, ең бастысы клеткада энергия трансформациям механизмдерін қамтамасыз етуші ферменттердің синтез процестерін ынталандырады.
- тиреоидты эсерді іске асырудың екінші механизмы 3,5 АМФ синтезін ынталандыру арқылы жэне ары қарай цАМФарқылы энергетикалык алмасудың негізгі ферменттерінің белсенділігін ынталандыру арқылы жүзеге асырылады.
Сонымен, тиреоидты гормондар өзінің физиологиялық эсерін корсете отыра, тыныс алу жэне басқа да митохондриальды ферменттердің санын, белсенділігін реттеу арқылы биологиялық тотыгу жэне тотыгу фосфорильдену процестерін ынталандыруды қамтамасыз етеді. Энергетикалық алмасуды тиреоидты реттеу механизмында ең бастысы, қалқанша безінің иодтирониндері тотығу фосфорильденудің терминальды реакциялар деңгейінде емес, тотыгушы субстраттардың энергиясын трансформациялау процесінің бүрынырақ бөлігінде эсер етеді- иондар тасымалдануына жэне баска да процестерге мембраналық потенциал энергиясын қалыптастыру жэне жеткізу деңгейінде, бүл эсер етуші мембраналардың омдық өткізгіштігінің өзгеруі есебінен жэне митохондриялардың тыныс алу тізбегінде ферменттік топтардың синтезінің индукциясы мен қүрылымдық жэне функционалдық ұйымдастығының қалыпты жагдайда болуы есебінен қамтамасыз етіледі
Агзада тиреоидты гормондардың деңгейі артқан кезде митохондриялардың тыныс алу тізбегінде тотыгушы субстраттар энергиясының протонды электрохимиялық мембраналық потенциалға трансформациялану процесі жылдамдайды, ол энергия жылу, электр энергиясына трансформациялануга, осмостық, механикалық жэне басқа қызметтерді атқару үшін немесе клеткалардың өзгешелік қызметтерінің орындалуы үшін өте ыңгайлы.
АТФ синтезі механизмының элсіреуі тыныс алу тізбегі бойынша электрондарды тасымалдау процесінің зақымдалуы есебінен гана емес, АТФ-синтетазалық реакция үшін энергияны жеткізуді шектеу есебінен де болады. Қалқанша безі гормондары іске қосатын бүл молекулярлы биоэнергетикалық процестердің барлығы да клеткалардың пролиферация-сымен, олардың өсуі жэне дифференцировкасымен, ал бүтін ағза деңгейінде тканьдар мен мүшелердің қалыпты өсуі жэне дамуы арқылы көрінеді. Зерттеулер көрсеткендей (Тапбергенов CO. 1989,1991,1994,1995), гипоталамо-гипофизарлы-тиреоидты ось симпато-адреналды жүйемен бірыңғай функционалды реттеуші комплекс - адрено-тиреоидты жүйені құрайды, оның ең басты қызметі - адаптация механизмдерін трансбиоэнергетикалық қамтамасыз ету болып табылады.
Адрено-тиреоидты жүйе - бірыңғай функциональды реттеуші комплекс болып табылады, оның басты компоненттері- гормон- медиаторлары бар симпато-адреналды жүйе жэне иодтирониндері бар қалқанша безі. Ағзаға кез-келген жедел немесе созылмалы стресс жағдайы, соның ішінде радиациялық эсер ету қалқанша безінің жэне симпато-адреналды жүйенің қызметінің зақымдалуын тудырады. Адрено-тиреоидты жүйе энергетикалық алмасудың жэне бақылауда болатын стресс- реакциялардың негізгі реттеуші жүйесі болып табылады. Ағзаның клеткалары, тканьдері жэне мүшелерінің қызметтерінің энергиямен қамтамасыз етілуінің зақымдалуын адрено-тиреоидты жүйені қүраушы компоненттерінің дисбалансы мен элсіреуін тудыратын кез-келген жагдайда байқауға болады. Ағзаның бақылаусыз стресс-процестерінің бақылаулы стресс реакцияларынан артық болу жағдайы адаптациялық механизмдердің бұзылуына экеледі жэне симпато-адреналды жүйенің ме-диаторлы бөлігінің элсіреуіне жэне адренотиреоидты жүйенің дисфункциясына байланысты болады. Адаптациялық механизмдердің бұзылуының қалпына келуі адрено-тиреоидты жүйенің қызметін қалпына келтіру арқылы жүзеге асырылады. Адаптациялық механизмдерді күшейту үшін жэне бақылаулы, бақылаусыз стресс- реакциялардың қалыптасуы үшін ағзада адаптациялық процестердің табиғи биореттеушілерінің- тиреоидты гормондар мен медиаторларының деңгейін қалпына келтіру қажет.
Cay адамның ағзасында Т4-62-141нмоль/л мөлшерінде, ал трийодтиронин (ТЗ)- 1.2-2.5 нмоль/л аралыгында болады. Қалканша безі йодтирониндерінің мөлшерден тыс болуы тиреотоксикозға экеледі (біріншілік диффузды тиреотоксикоз- Базед ауруы). Мүндай аурулардың қанында ТЗ көп болады.Тиреотоксикоздың белгілері- көмірсулар мен липидтердің жедел ыдырауы, олардың митохондрияларда тез тотығуы болып табылады. Тканьдік тыныс алу жеделдейді, оттегіні пайдалану артады. Митохондриялар көлемі мен формасын өзгертеді, ісінеді. Сол себептен тиреотоксикозды "митохондриялар ауруы" деп те атайды. Біртіндеп белоктың синтезінен белоктың ыдырау процестері басым бола бастайды. Базед ауруының сыртқы белгілері негізгі алмасудың көбеюімен, дененің температурасының жоғарылауымен, дене салмагының азаюымен, ерекше қозғыштықпен, үйқы қашуымен, тахикардиямен,экзофтальмен сипатталады.
Бүйрекүсті безінің милы затының гормондары, химиялық табиғаты. Биосинтезі, зат алмасуға және биоэнергетикалық процестерге әсері.
Бүйрекүсті безінің милы затыныңгормондары – биогенді моноаминдерге жататын физиологиялық белсенді заттар. Оларда пирокатехин ядросы бар, сондыктан оларды катехоламиндер деп атайды. Бүйрек үсті безінің милы затының хромаффинды клеткаларында дофамин түзілу кезеңі арқылы тирозиннен норадреналин жэне адреналин гормондары синтезделеді.
Бүйрек үсті безінің милы затының гормондарының тирозиннен биосинтезі:

Тирозиннен түзілетін катехоламиндердің қатарында қасиеті мен қызметшщ модуляторлықтан (дофамин) медиаторлыққа (норадреналин) және гормоналдыққа (адреналин) ауысуы байқалады. Бүл жағдайда осы қатардағы жеке өкілдердің қызметтерінің бірін-бірі жабуы байқалады: дофамин мен норадреналинның модуляторлық эсері, норадреналин мен адреналинның медиаторлық жэне гормоналдық эсері.
Дофаминның норадреналинге айналуын дофамин- бета- гидроксилаза ферменті катализдейді, ол эндогенді норадреналинмен қатар митохондриалды фракцияда табылған. Дофаминнің норадреналинге ферменттік айналуы үшін аскорбин қышқылының қатысуы қажет.
Адреналин - бүйрек үсті безінің хромаффинды клеткаларының цитозолінде орналасқан норадреналиннен фенил- этиламин-N- метилтрансфераза ферментінің қатысуымен синтезделеді. Гипофизэктемия, глюкокортикоидтар жетіспеушілігі кезінде бүл ферменттің синтезі бүзылады, ал АКТГ-ны жіберу адреналиннің түзілуін тудырады.
Дофамин мен норадреналин эртүрлі молшерде барлық гипоталамусты ядроларда кездесетіні белгілі. Ми құрылымында дофаминды рецепторлар бар, олар өз эсерін дофамин-ге сезімтал аденилатциклаза арқылы жүзеге асырады. Қоздырылуы гипертермияға экелетін адренергиялық рецепторларға қарағанда, дофаминды рецепторлар, сол сияқты дофаминның бөлінуін ынталандыратын заттар гипотермиялық эсер көрсетеді. Дофаминның аз концентрациясы (1-Змкг/кг) артериалды қысымды төмендетеді, ал жоғары концентрациясы (9-81 мкг/ кг) артериалды қысымды жогарылатады жэне миокардтың жиырылу күшін арттырады. Сонымен қатар, дофамин дофаминді рецепторларға эсерінен басқа, жүректің бета- адреноре-цепторларына жэне қан тамырлардың тарылуын тудыратын альфа-адренорецепторларга эсер етеді. Алайда, бүл жагдайда дофаминнен түзілетін норадреналинның да эсерін естен шығармау керек. Норадреналин сияқты, дофамин да тек жүрек- қан тамыр жүйесіне ғана эсер етіп қоймай, сонымен бірге көмірсулар жэне липидтер алмасуына да эсер етеді. ДОФА, дофамин, норадреналин мен адреналинның метаболитикалық эсерлерін салыстыру ДОФА-ға қарағанда дофаминның сүт жэне пирожүзім қышқылының жинақталуына көбірек эсер ететінін көрсетті.
Адреналин- бүйрек үсті безінің милы затының гормоны, ол күшті оксидо- редукторлы қасиеті бар қосылыс, симпатикалық жүйке жүйесінің тонусын арттырып, оның орталықтарын қоздырады.
Дофамин мен норадреналин эртүрлі молшерде барлық гипоталамусты ядроларда кездесетіні белгілі. Ми құрылымында дофаминды рецепторлар бар, олар өз эсерін дофамин-ге сезімтал аденилатциклаза арқылы жүзеге асырады. Қоздырылуы гипертермияға экелетін адренергиялық рецепторларға қарағанда, дофаминды рецепторлар, сол сияқты дофаминның бөлінуін ынталандыратын заттар гипотермиялық эсер көрсетеді. Дофаминның аз концентрациясы (1-Змкг/кг) артериалды қысымды төмендетеді, ал жоғары концентрациясы (9-81 мкг/ кг) артериалды қысымды жогарылатады жэне миокардтың жиырылу күшін арттырады. Сонымен қатар, дофамин дофаминді рецепторларға эсерінен басқа, жүректің бета- адреноре-цепторларына жэне қан тамырлардың тарылуын тудыратын альфа-адренорецепторларга эсер етеді. Алайда, бүл жагдайда дофаминнен түзілетін норадреналинның да эсерін естен шығармау керек. Норадреналин сияқты, дофамин да тек жүрек- қан тамыр жүйесіне ғана эсер етіп қоймай, сонымен бірге көмірсулар жэне липидтер алмасуына да эсер етеді. ДОФА, дофамин, норадреналин мен адреналинның метаболитикалық эсерлерін салыстыру ДОФА-ға қарағанда дофаминның сүт жэне пирожүзім қышқылының жинақталуына көбірек эсер ететінін көрсетті.
Жете физиологиялық функционалды жэне фармакологиялық сараптама негізінде Комиссаров И.В. адренорецепторлардың жэне соған байланысты катехоламиндердің функционалдық эсерінің жіктелуін үсынды:
- альфа 1- адренергиялық қозу жэне мүшелердің тегіс бүлшық етінің жиырылу рецепторлары.
- альфа 2- асқазан- ішек жолы мүшелерінің тегіс бұлшық етінің адренергиялық тежелуінің рецепторлары.
- бета- 1- катехоламиндердің жүрекке, асқазан- ішек жолының тегіс бұлшық еттеріне эсерін жеткізуші жэне катехоламиндердің липолитикалық эсерін қамтамасыз етуші рецепторлар.
- бета- 2- рецепторлар, олар арқылы вазодилятация, жатырдың тегіс бүлшық етінің тонусының төмендеуі жүреді жэне қаңқа бүлшық еттерінде гликогенолиз ынталандыры-лады.
- бауырдағы гликогенолиз жэне липолиздың гамма- рецепторлары.
Сонымен, тканьдерде катехоламиндерге деген қосынды метаболитикалық реакция энергетикалык субстраттардың жогары концентрациясын қамтамасыз етуге экеледі: бауырдан-глюкозаның, май тканінен- бос май қышқылдарыньщ. Белокты алмасуға катехоламиндердің эсері гликоген қоры таусылған жагдайда немесе стресс жағдайында байқалатын адреналинның коп молшерде бөлінуі кезінде байқалады.
Ағзаның кез-келген эффекторлы клеткасында катехоламиндердің барлық физиологиялық қызметтерінің іске асырылу механизміндегі ең бастысы- олардың энергетикалык алмасуды реттеуге қатысуы болып табылады. Азотты, көмірсулы, липидті алмасудагы катехоламиндер эсерінен болатын өзгерістер клеткадагы биоэнергетиканы қамтамасыз етуге бағытталған.
Калоригенді эсер ету катехоламиндердің негізгі эсері болып табылатыны белгілі. Катехоламиндердің калоригенді эсері тканьдердің оттегіні пайдалануының күшеюімен катар жүреді, ол биологиялық тотыгу мен тканьдік тыныс алу процестерін катехоламиндер ынталандыратынын көрсетеді. Катехоламиндердің калоригенді эсері жүректе бос май кышкылдарының тотыгуының күшеюімен, бауырда жэне басқа мүшелерде көмірсулар мен липидтерді еселене пайдалануымен байланысты. Суыққа төзімді жануарларга жіберілген норадреналинның калоригенді эсерінің күшеюі қаңқа бүлшық еттері мен қоңыр майдың функционалды белсенділігінің өзгеруімен байланысты жэне адреналин ынталандыратын оттегіні тұтынуымен байланысты.
Ағзаға адреналинды енгізу оттегіні пайдалану мен макроэргтер мөлшерінің деңгейінің сэйкессіздігін тудырады. Адреналинге қарағанда норадреналин элсіз түрде АТФ-тың деңгейін төмендетеді де, АТФ/АДФ қатынасы да төмендейді.
Катехоламиндердің жүрек бұлшық еті жиырылуына негізгі әсерлері:
Саркоплазмаға Са++ кіруін, жиырылу күшін жоғарылату. 2. Аденилатциклазаның белсенуін, тропонин фосфорильденуінің жылдамдығының, жиырылу жылдамдығының жоғарылауы. 3. Жиырылуды энергиямен қамт. етудің белсенуі, энергия алмасу жылдамдауы.

