




Пущино - 2016
Доклад
Редокс-статус клетки, окислительный стресс и митохондрии
Основные сокращения
АФК – активные формы кислорода
ИИ – ионизирующее излучение
ПОЛ – перекисное окисление липидов
СОД – супероксиддисмутаза
ADP — аденозиндифосфат;
AIF — апоптозин- дуцирующий фактор;
AP-1 — белок-активатор 1;
ARTs — (ADP-Rib)трансферазы;
ATP — аденозинтрифосфат;
cADP-ри- боза — циклическая ADP-рибоза;
Его1 — оксидоредуктаза 1 эндоплазматического ретикулума;
FAD/FADH2 — флавинаденин- динуклеотид окисленный/восстановленный;
FMN/FMNH2 — флавинмононуклеотид окисленный/восстановленный;
GPx — глутатионпероксидаза;
GR — глутатионредуктаза;
Grx — глутаредоксин;
GSSG/2GSH — глутатион окисленный (глутатиондисульфидУвосстановленный;
IP3R— рецептор инозитол-1,4,5-трисфосфата;
MPT —неспецифическая проницаемость митохондрий;
NAADP — адениндинуклеотид- фосфат никотиновой кислоты;
NAD+/NADH — Р-никотин- амидадениндинуклеотид
АФК - активные формы кислорода;
ДЛДГ - дигид- ролипоамиддегидрогеназа;
ДОДГ - дигидрооротатдегидрогеназа;
а-КГДГК - а-кетоглутаратдегидрогеназный комплекс;
МАО - моноаминооксидаза;
мГФДГ - митохондриальная а-глицерофосфатдегидрогеназа;
ПДГК - пиру- ватдегидрогеназный комплекс;
СМЧ - субмитохондриальные частицы;
СОД - супероксиддисмутаза;
ЭПФ - электронпереносящий флавопротеин;
Amplex Red - 10-ацетил-3,7-дигидрофеноксазин;
NOX - NAD(P)H-оксидаза;
Др – разность электрохимических потенциалов ионов водорода.
GPX — глутатионпероксидаза
F!FQATФ-синтетаза — крупный ферментный комплекс, образуемый девятью полипептидными цепями. Пять из них составляют сферическую головку ком-плекса, называемую F, -АТФ-си нтстазой. где и происходит синтез АТФ. Четыре оставшиеся полипептид- ные цепи формируют гидрофобную субструктуру F0, встроенную во внутреннюю митохондриальную мембрану, и представляют собой «канал», через который протоны переходят к активному центру АТФ- синтетазы, т.е. к Ft.
ND — субъединицы фермента NADH-дегидрогеназы
Ьр — пара азотистых оснований (base pair)
КБ — тысяча пар оснований (kilobase)
Транзиция — замена одного пиримидина на другой пиримидин или одного пурина на другой пурин
Трансверсия — замена пиримидина на пурин или, наоборот, пурина на пиримидин
Турновер — постоянное самообновление предсу- ществующих субклеточных структур, макромолекул и их комплексов путем распада и последующей замены их на синтезированные de novo аналоги
Процессинг — высокоспецифичное расщепление крупного транскрипта — предшественника — на входящие в его состав функционально активные молеку-лы РНК
Wobble позиция — «качание» или неоднозначное соответствие третьего основания антикодона. Если первые два основания антикодона спариваются с соответствующими основаниями кодона строго по правилу Уотсона—Крика, то в третьем положении допускается нарушение этого правила комплементарности, что является одной из причин вырожденности генетического кода.
Содержание
Основные сокращения.. 2
Содержание. 5
Ведение. Редокс-статус клетки. 7
Активные формы кислорода (АФК) 8
Синглетный кислород. 12
Пероксид водорода. 13
Супероксид-анион радикал. 14
Гидроксил-радикал. 16
Окислительный стресс. 18
Окислительная модификация липидов. ПОЛ. 21
Окислительная модификация белков. ДЖРБ. 23
Окислительная модификация нуклеиновых кислот. Повреждение ДНК. 35
Антиоксидантная и прооксидантная системы организма\. 37
Основные редокс-пары клетки. 38
GSSG/2GSH.. 39
TrxSS/Trx(SH)2 45
NAD+/NADH.. 47
NADP+/NADPH.. 53
ФЛАВИНЫ.. 56
Прооксиданты.. 59
NAD(P)H-оксидазы.. 60
Антиоксиданты.. 61
Ферментативная АОС. 61
Другие антиоксиданты.. 65
Митохондрии, их строение и функции. 69
Цикл Кребса, ЭТЦ и АФК. 72
НАD(Р)Н-ОКСИДАЗА (NOX4) 78
Моноаминооксидаза. 79
Ацил-СоА дегидрогеназа, электронпереносящий флавопротеин и его дегидрогеназа. 80
Дыхательный комплекс I 81
Комплекс II 90
Комплекс III 92
Дигидролипоамиддегидрогеназа. 96
Митохондриальный геном. Повреждение митохондриального генома и митохондриальные болезни. 98
Мутации митохондриальной ДНК. 102
Митохондриальная дисфункция. 108
Митохондриальные болезни. 109
Физиологическое значение генерации АФК митохондриями. 110
Митохондрии, апоптоз и АФК. 114
Окислительный стресс при онкологии.. 119
Радиационно-индуцированный окислительный стресс. 120
Хемилюминисценция.. 122
Литература.. 122
Ведение. Редокс-статус клетки.
Редокс-потенциал характеризует активность восстановителей или окислителей любого раствора или способность этого раствора отдавать или принимать электроны. Цитозоль каждой клетки нашего организма – это раствор, где протекают сложнейшие биохимические реакции, в т. ч. и окислительно-восстановительные. Баланс окислительно-восстановительных процессов в клетке определяет клеточный редокс-статус (от англ. redox — reduction-oxidation reaction). Все формы жизни сохраняют восстанавливающую среду внутри своих клеток. Когда редокс статус клетки увеличивается настолько, что клетка не может его скомпенисировать начинается окислительный стресс.
Клеточный «редокс-статус» поддерживается специализированными ферментами в результате постоянного притока энергии. Аэробная энергетика всех живых существ обеспечивается свободной энергией окислительно-восстановительной реакции между восстановителем (внутриклеточная среда, усредненный редокс потенциал которой составляет примерно -320 мВ) и окислителем - кислородом, редокс- потенциал которого в реакции образования воды при нейтральных значениях рН равен примерно +800 мВ. Кислород - «сильный» окислитель в том смысле, что реакция образования воды:
О2 + 4 е + 4 Н+ = 2 Н2О (1)
при восстановлении кислорода электронами, поставляемыми восстановительными эквивалентами клетки (главным образом NADH), термодинамически необратима. С другой стороны, кислород - «плохой» окислитель. Электронная структура его молекулы (два неспаренных электрона с одинаковыми спиновыми числами на разных п-орбиталях) такова, что кислород не может принять одновременно два электрона от подавляющего числа органических молекул - доноров пары электронов с антипараллельными спинами. Кинетическая инертность кислорода обеспечивает устойчивость многочисленных органических молекул в окружающей нас среде, содержащей 21% газообразного кислорода. Спиновый запрет реакций окисления органических соединений кислородом в сочетании с термодинамической необратимостью таких реакций и послужил физико-химической основой того, что кислород выбран эволюцией в качестве окислителя. Эволюция решила проблему кинетической инертности кислорода, создав ферменты- оксидазы, катализирующие восстановление кислорода до воды (цитохромоксидазы, реакция 1), или до перекиси водорода (другие оксидазы, реакция 2):
О2 + 2 е + 2 Н+ = Н2О2. (2)
Проблема спинового запрета решается этими ферментами так, что восстановление кислорода в суммарных реакциях (1) или (2) происходит ступенчато с помощью одноэлектронных доноров-кофакторов: ионов металлов переменной валентности (железо, марганец, молибден, медь) и (или) органических молекул, способных существовать в свободнорадикальном состоянии (флавины, хиноны).
На промежуточных этапах этого процесса образуются АФК, которые, в отличии от собственно триплетного кислорода, кинетической инертности не имеют и в избыточном количестве могут вызвать нарушение редокс-статуса клетки. Поддержание клеточного редокс-статуса играет важную роль в таких процессах как синтез ДНК, экспрессия генов, ферментативная активность. Изменения редокс-состояния молекул, являющиеся следствием стрессовых воздействий или результатом активности самих клеток, вовлечены в редокс-регуляцию клеточных процессов.
Активные формы кислорода (АФК)
Активные формы кислорода (англ. ROS, Reactive Oxygen Species) – это высокореакционные, преимущественно радикальные кислородные соединения, образующиеся в живых организмах в результате неполного восстановления молекулярного кислорода или изменения спина одного из его электронов, находящихся на внешних орбиталях. В настоящее время для объединения широкого класса кислородсодержащих соединений радикальной и нерадикальной природы так же используется термин “активированные кислородные метаболиты” (АКМ).
Под термином «активные формы кислорода» в различных источниках объединяют: супероксидный анион-радикал O2-, пероксид водорода HOOH, гидроксильный радикал ОН•, гидроперекисный радикал НО2•, синглетный кислород 1O2, озон O3, алкоксильные RO• и пероксильные радикалы ROO-, гипогалоиды HOCl, HOBr, HOI, оксид азота NO•, гидропероксил радикал НОО, пероксинитрит ONOO- и ряд других соединений.
АФК часто называют свободными радикалами, но это не совсем корректно: свободные радикалы — это высокоактивные молекулы или атомы, имеющие один или несколько неспаренных электронов на внешней орбитали, что делает их особенно активными и «агрессивными». Свободные радикалы стремятся вернуть себе недостающий электрон, отняв его от окружающих молекул. Такие соединения как HOOH и 1O2 свободными радикалами не являются, но при этом они однозначно относятся к АФК. Наиболее изученными и представляющими наибольший интерес АФК в клетке остаются супероксидный анион-радикал O2, пероксид водорода HOOH и гидроксильный радикал ОН•, а также оксид азота и перокси нитрит, которые ряд авторов относя к активным фомам азота.
Основные свойства активных форм кислорода: высокая реакционная способность, короткое время жизни, малый или относительно малый радиус диффузии и относительно низкая концентрация в тканях (таблица, слайд).
Свободнорадикальные реакции могут быть как ферментативной, так и неферментативной природы. К первым относятся реакции дыхательной цепи, синтеза простагландинов, цитохрома Р-450, фагоцитоза и др.; ко вторым - катализируемые Cu и Fe процессы окисления органических соединений, реакции, индуцируемые ионизирующим излучением, ультрафиолетовым, и др. Образование АФК происходит постоянно и в процессе нормальной жизнедеятельности всех аэробных организмов.
Различные источники АФК можно разделить на две большие группы (Frenkel, 1992):
| Эндогенные источники: | Экзогенные источники: |
| 1. Стимуляция фагоцитирующих клеток. 2. Нефагоцитирующие клетки. 3. Редокс-цикл “ хинон-семихинон”. 4. Индукция синтеза прооксидантных ферментов. 5. Ингибирование антиоксидантных ферментов. 6. Индукция синтеза СоА - оксидаз жирных кислот. 7. Ишемия и реперфузия. | 1. Ионизирующее излучение. 2. УФ - излучение. 3. Воздействие ксенобиотиков. 4. Озон. |
Синглетный кислород
В основном состоянии кислород триплетен, спин равен 1, тогда как его вышележащие возбужденные состояния синглетны (спин равен 0) (рис. слайд). Наинижайшее состояние электронного возбуждения называют синглетным кислородом (1O2). Cинглетным называют молекурный кислород в состоянии наименьшего электронного возбуждения. Изменение спина одного из электронов, находящихся на π-орбиталях в молекуле кислорода, приводит к образованию возбужденного синглетного состояния, энергия которого на 96,3 кДж/моль больше энергии основного триплетного состояния.
Благодаря высокой реакционной способности, синглетный кислород вступает в окислительные реакции с различными органическими соединениями: принимает участие в повреждении нуклеиновых кислот и канцерогенезе, в инициировании ПОЛ и в возникновении биохемилюминесцеции, ингибирует Са2+-АТФазу. Кроме того, синглетный кислород агрессивен в отношении молекул с двойной связью. Конечным продуктом таких реакций обычно является образование гидроперекисей органических молекул – один из важнейших в процессах перекисного окисления ненасыщенных липидов в биомембранах.
Синглетный кислород как сопутствующий продукт образуется при многих ферментативных реакциях, катализируемых каталазой, миелопероксидазой и т.п. Основное биологическое значение синглетного кислорода становится существенным при различных фотоиндуцированных процессах, в частности – фотосенсибилизации.
Пероксид водорода
Замечательное свойство большинства ферментов-оксидаз состоит в том, что они катализируют ступенчатое восстановление кислорода до воды или перекиси водорода, не освобождая в окружающую среду промежуточно восстановленных реакционноспособных и тем самым потенциально опасных для клеток форм кислорода. Только очень небольшая часть потребляемого кислорода восстанавливается до перекиси водорода (реакция 2), которая, в свою очередь, сама служит субстратом-окислителем для так называемых пероксидаз:
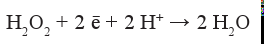
или восстановителем и окислителем в реакции её дисмутации (каталаза):
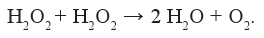
В клетке пероксид водорода может образовываться посредством преобразования супероксидного анион-радикала как спонтанно, так и при каталитическом участии супероксиддисмутазы, а также в процессе прямого синтеза в реакциях с участием гликолатоксидаз или оксидаз D-аминокислот в пероксисомах. В митохондриях описаны источники образования пероксида водорода - окисление сукцината и NADH (Комплексы I и II).
Пероксид водорода представляет большой интерес исследователей, поскольку выделяется среди других активных форм кислорода не высокой окислительной способностью, но большой продолжительностью жизни. Сам пероксид водорода не обладает свойствами сильного окислителя, однако легко преобразуется в другие, более агрессивные молекулы. Пероксид водорода может выступать источником образования ОН•в присутствии двухвалентного железа или превращаться в гипохлорит-анион (ОС1-) ферментом миелопероксидазой.
Перекись водорода относится к окислителям средней силы, при этом, не будучи радикалом, она взаимодействует с веществами как радикальным, так и нерадикальным путем. Пероксид водорода может проявлять сосудосуживающий и цитотоксический эффекты, он участвует в подавлении пролиферации лимфоцитов. Уровень Н2О2 удаляется в основном каталазой, а в митохондриях и эндоплазматическом ретикулуме с помощью глутатионпероксидазы.
Показано, что Н2О2 вызывает гибель опухолевых клеток, эндотелиоцитов, NK-клеток и Т-лимфоцитов ВИЧ-инфицированных людей посредством апоптоза. На уровне целого организма отравление Н2О2 приводит к обширной газовой эмболии и повышению активности ядер блуждающего нерва; в результате нарушается функция дыхания и наступает смерть.
Пероксид водорода в малых концентрациях выполняет ряд важных функций в клетке. Так, он используется фагоцитами в реакции, катализируемой миелопероксидазой, для образования гипохлорита, который, в свою очередь, обладает свойством разрушать клеточную стенку бактерий, тем самым защищая организм от чужеродной клетки. Однако кроме защитной функции большое внимание уделяется важной роли пероксида водорода в регуляции клеточной пролиферации и апоптоза. Несмотря на слабые окислительные свойства, он способен окислять SH- группы белков, вызывая тем самым модификацию рецепторных белков, имитируя функцию лиганда. Таким образом, он принимает участие в возникновении и передаче клеточных сигналов.
Супероксид-анион радикал
Существование каталитически активных ионов металлов переменной валентности и (или) органических соединений, способных к одноэлектронным реакциям, в составе оксидоредуктаз или белках-переносчиках кислорода (миоглобин, гемоглобин) приводит к потенциальной возможности одноэлектронного восстановления кислорода до супероксид-радикала - аниона при физиологических значениях рН (pKa супероксида равен 4,7):
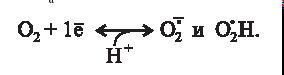
Супероксид - анион радикал при взаимодействии с протоном приводит к образованию гидроперекисного радикала (НО2-•). При кислых значениях рН НО2-•, так же, как и Н2О2, не имеет заряда и может проходить через клеточную мембрану.
Супероксид в водных растворах при рН > 5 (см. рКа в уравнении 5) нестабилен и достаточно быстро неферментативно дисмутирует:
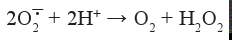
Основным источником образования супероксиданион радикала в биологических системах являются НАДФ-оксидаза, ксантиноксидаза, митохондриальная цитохром-с-оксидаза и микросомальные миелооксидазы. Наибольшей способностью продуцировать О2-• у человека и животных обладают фагоцитирующие клетки
В живых организмах существует группа специализированных ферментов, отвечающих за снижение уровня супероксид-анион радикала – супероксиддисмутаза (СОД). Генерация О2-• играет важную роль в реализации антимикробного, цитотоксического и иммунорегуляторного действия фагоцитов. Супероксид-анион радикал может вызвать окислительную модификацию липопротеинов сыворотки и нуклеотидов.
Супероксид-анион радикал принимает участие в многочисленных реакциях с образованием различных реактивных соединений, таких как пероксид водорода, пероксинитрит, окисленный липопротеин низкой плотности или гипогалоиды.
В результате активации под действием О2-• NO-синтазы в тканях образуется NO- радикал, обладающий свойствами вторичного мессенджера и является эндотелиальным фактором расширения сосудов. Вместе с тем, супероксиданионрадикал может уменьшать уровень NO• - радикала за счет превращения его в пероксинитрит. Последний может способствовать индукции апоптоза, а в ходе своего спонтанного распада превращаться в ОН•.
Гидроксил-радикал
Одноэлектронное восстановление перекиси водорода (третий электрон на пути ступенчатого четырех-электронного восстановления молекулярного кислорода до воды) приводит к возникновению крайне реакционноспособного (термодинамически и кинетически) гидроксил-радикала:
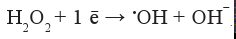
Донором для одноэлектронного восстановления перекиси водорода может служить супероксид-радикал в реакции, катализируемой металлами переменной валентности (Fe2+, Cu+, Co2+, Mn2+, V2+, Cr4+):
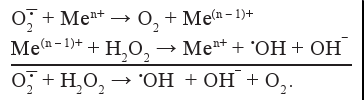
Рекомбинация Н2О2 и Fe2+, лежащая в основе реакции Фентона, является главным механизмом образования ОН•. При взаимодействии Н2О2 и О2-• (реакция Хабера-Вайса) в присутствии металлов с переменной валентностью также образуется ОН•.
Гидроксильный радикал является наиболее реакционноспособным радикалом среди АФК. Основные типы повреждений биомолекул гидроксильными радикалами - это отрыв атома водорода; присоединение к молекулам по двойным связям; отрыв электрона (редко). Кроме того гидроксильные радикалы способны разорвать любую С–Н связь, что зачастую приводит к повреждениям структуры белков и нуклеиновых кислот. Особенно активно он взаимодействует с теми липидами, жирнокислотные радикалы которых содержат ненасыщенные двойные связи; это приводит к образованию липидных гидроперекисей в ходе цепных реакций (так называемое перекисное окисление липидов - ПОЛ). для его элиминации в клетке не существует специализированных ферментных систем. Считается, что цитотоксическое действие кислородных радикалов более чем на 50% обусловлено ОН-радикалами, при этом в клетках выделяют два критических объекта повреждения: нуклеиновые кислоты и мембранные белки.
В нормальных условиях АФК поступают в организм с кислородом воздуха, потребляемыми жидкостями и образуются в функциональных системах его клеток в процессе синтеза простангландинов, в результате респираторного взрыва фагоцитирующих клеток, при восстановлении кислорода в дыхательной цепи митохондрии, а также при окислении ксенобиотиков и эндогенных субстратов в митохондриальной цепи транспорта электронов. При этом определенное количество АФК представляет собой постоянный окислительный «фон», присутствующий в живых организмах сигнализирующий о постоянстве окислительно-восстановительного гомеостаза в клетке (редокс-статуса).
Окислительный стресс
Все клеточные процессы характеризуются, тем, что протекают в условиях окислительно-восстановительного гомеостаза, обусловленного изменением в окислительно-восстановительных реакциях множества веществ. Дисбаланс окислительно-восстановительного гомейостаза внутри клетки или ткани, за частую инициируется и/или сопроваждается увеличением концентрации АФК. В норме активные формы кислорода образуются в физиологических концентрациях и играют важную сигнальную и сигнально-регуляторную роль. Избыточную генерацию АФК свыше уровня антиоксидантной защиты, сопровождающуюся повреждением клеточного содержимого, называют окислительным стрессом.
Причинами окислительного стресса называют как внутренние факторы, такие как недостаточная активность антиоксидантных ферментов по сравнению с прооксидантными, так и внешние - ионизирующее и ультрафиолетовое излучение, лекарственные препараты, ксенобиотики.
По концентрации окислительных агентов и реакции на них клеток окислительный стресс условно подразделяют на уровни:
1. Низкий уровень - клетка обеспечивает себе достаточную защиту. При этом изменения наблюдаются большей частью в синтезе белков. В норме клетки могут длительное время пребывать в состоянии непрерывного окислительного стресса низкого уровня без серьезных последствий.
2. Средний уровень - клетка осуществляет адаптацию к стрессу.
3. Высокий уровень - клетка переходит в состояние выживания, клеточный рост и деление в этом случае практически останавливаются. Дальнейшее повышение концентрации активных форм кислорода приводит к массовой гибели клеток.
Очевидно, что опасность для жизни клетки возникает при достижении высокого уровня окислительного стресса, однако отмечено повышение риска становления и развития злокачественной опухоли под воздействием окислительного стресса низкого и среднего уровня (рис. 2).
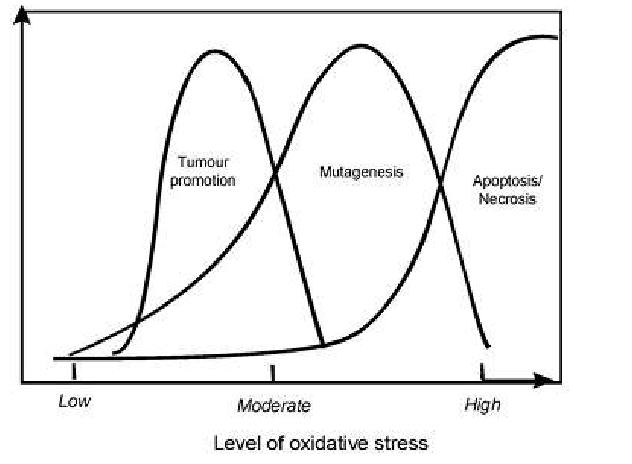
Рис. 2. Дозозависимый эффект взаимосвязи между уровнем окислительного стресса и процессом прогрессирования опухоли, процессом мутагенеза и процессом апоптоза/некроза.
Такие ткани организма как сетчатка, легкие, мозг обладают наибольшей чувствительностью к окислительному стрессу, что связано с особенностями их метаболизма и морфологией. Повышенной же устойчивостью отличаются клетки кожи, вероятно в силу их постоянного нахождения в прямом контакте с кислородом воздуха.
Окислительный стресс с накоплением в тканях и биологических жидкостях АФК и вторичных продуктов окислительной модификации макромолекул обнаружен при более чем 60 болезнях и синдромах. У человека окислительный стресс может стать первопричиной или неотъемлемой составляющей многих серьёзных заболеваний, таких как атеросклероз, гипертензия, болезнь Альцгеймера, диабет, а также одной из основных составляющих процесса старения. Количество АФК увеличивается при любом патологическом процессе прямо пропорционально тяжести состояния. Такое многообразие негативных эффектов на уровне целого организма на клеточном уровне обуславливается тремя основными процессами:
· окислительная модификация липидов;
· окислительная модификация белков;
· окислительная модификация нуклеиновых кислот.

