




Фракціонування і визначення окремих амінокислот. Деякі амінокислоти дають кольорові реакції або володіють характерним спектром поглинання, що створює можливість їх кількісного визначення. Існують, проте, і більш загальні методи, що дозволяють ідентифікувати, а потім і визначити вміст кожної з амінокислот в суміші після фракціонування. Ці методи можуть бути використані при дослідженні тканинних екстрактів, біологічних рідин (плазми крові, сечі) або білкових гідролізатів, в яких зв'язок між амінокислотами вже розірваний.
Хроматографія на папері (Мартін, Гордон і Синдж). Цей метод відіграє важливу роль при аналізі амінокислот, і сама техніка хроматографії на папері була розроблена саме для цієї мети.
Коли мають справу з складною сумішшю амінокислот, наприклад при аналізі гідролізата білків, доводиться вдаватися до двомірної хроматографії. При проведені хроматографії в одному напрямі часто використовують як розчинник фенол, а в перпендикулярному – суміш бутанола і водного розчину оцтової кислоти. Амінокислоти проявляють за допомогою нінгідрина при підвищеній температурі. Амінокислоти, які відносяться до однієї групи, розташовуються на одній плавній кривій.
Іонообмінна хроматографія на колонці (Мур і Стейн). Цей метод дозволяє проводити кількісне визначення амінокислот у суміші. Скляну колонку заповнюють смолою, яка містить на матриці полістиролу угрупування – SO3H-Na+. В кислому середовищі амінокислоти поводять себе як основи. Вони утворюють катіони, здатні витісняти частину іонів Na+ і, займаючи їх місце, затримуватися на колонці. Чим більш основною є дана амінокислота, тим міцніше вона зв'язується електростатичними силами із смолою. Потім через колонку пропускають елюючу рідину, поступово збільшуючи її рН (градієнт рН). Коли рН елюента наближається до значення рН, відповідного даній амінокислоті, вона відділяється від смоли і починає рухатися уздовж колонки. Оскільки величини pHi для різних амінокислот неоднакові, амінокислоти виходять з колонки по черзі. Необхідно, проте, мати на увазі, що міцність зв'язку між амінокислотою і смолою обумовлена не тільки електростатичними силами, а залежить також від сил іншої природи. Тому в деяких випадках порядок виходу амінокислот може не відповідати порядку зміни величини рНi. Процес проведення хроматографії залежить від температури. Методика колоночної хроматографії використовується в двох варіантах:
а) в поєднанні з колектором фракцій. Амінокислоти збирають в окремі пробірки у вигляді невеликих фракцій. В кожну з пробірок додають нінгідрин і нагрівають (реакцію з нінгідрином проводять зазвичай при підвищеній температурі). За допомогою спектрофотометра вимірюють потім спектри поглинання кожної з фракцій і, виходячи з отриманих даних, обчислюють концентрацію кожної амінокислоти. Таким чином одержують серію кривих, що відповідають послідовному виходу амінокислот. Природу кожної амінокислоти встановлюють за об'ємом елюента, що витрачається на зняття цієї амінокислоти із смоли, з урахуванням заздалегідь проведених калібрувальних дослідів;
б) з автоматичним аналізатором, що працює за таким принципом. Рідина, що витікає з колонки, за допомогою насоса змішується з нінгідрином. Суміш проходить спочатку через водяну баню, де підігрівається, а потім через кювету спектрофотометра, який безперервно реєструє поглинання світла при 570 нм (фіолетове забарвлення нінгідрииа зі всіма амінокислотами) і при 440 нм (жовте забарвлення проліна з нінгідрином). Дані спектрофотометра реєструються на стрічці, що рухається, або на листі паперу. Площа, яка обмежена кривою і базовою лінією, пропорційна концентрації відповідної амінокислоти.
Електрофорез. Завдяки своєму електричному заряду амінокислоти можуть переміщуватися в електричному полі (рис. 1). Проте через малий розмір молекул вони легко дифундують і тому їх розділення можливе тільки при використанні твердої основи, наприклад паперу або крохмалю. Розділення йде по групах. Так, наприклад, при рН 4,0 дикарбонові амінокислоти ще залишаються кислотами і аніонами. Вони прямують до позитивного полюса – аноду. Нейтральні ж і лужні амінокислоти рухаються в цих умовах до негативного полюса – катоду, оскільки при рН 4,0 вони несуть позитивний заряд.
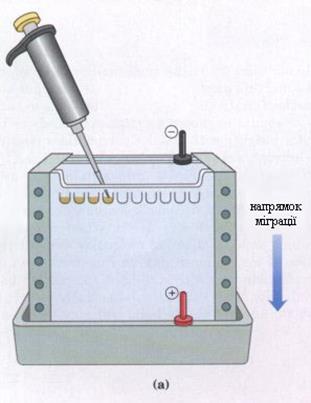
Рис. 1. Прилад для проведення електрофорезу (а), електрофореграма (b).
Будова білків
Всі білки являються високомолекулярними поліпептидами. Умовну границю між великими поліпептидами і білками проводять в області молекулярних мас 8000 – 10000.
Характеристика зв’язків амінокислот у молекулах білка. Для нативних білків, що мають специфічні фізико-хімічні та біологічні властивості, характерним є наявність нижчих та вищих рівнів структури. Для кожного рівня структури характерні певні види зв’язків, за рахунок яких відбувається надійна стабілізація білкових молекул.
Припущення відносно сил, що забезпечують утворення стабільних структур, було висловлене Д.Л. Талмудом та С.Є. Бреслером у 1944 p. Наступні дослідження дали змогу на основі цих припущень сформулювати загальні положення про сили, що стабілізують білкові молекули. Згідно з даними положеннями структура білків є результатом дії таких сил: пептидних зв’язків між CO–NH–групами амінокислот; дисульфідних зв’язків, які утворюються між залишками амінокислоти цистеїну; водневих зв’язків між киснем карбонільної і воднем імінної груп пептидних угруповань; гідрофобної взаємодії.
Отже, стабілізація структури білкової молекули забезпечується за рахунок ковалентних зв’язків та сил слабкої взаємодії. При цьому в стабілізації кожного виду структури вирішальна роль належить одному із зв’язків, а решта відіграють допоміжну роль. Так, ковалентні зв’язки (сили міцної взаємодії) забезпечують стабілізацію первинної і третинної структур (пептидний та дисульфідний зв’язки). Решта видів структури стабілізується за рахунок сил слабкої взаємодії водневих та іонних зв’язків, сил Ван-дер-Ваальса, гідрофобної взаємодії тощо.
Пептидний зв’язок. Коли карбоксильна і аміногрупа з’єднуються, утворюючи пептидний зв’язок, амінокислоти перетворюються в амінокислотні залишки, утворюючи пептид. Схематично даний процес можна записати так:
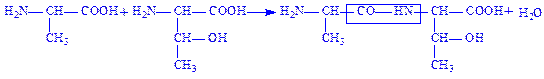
Пептид складається із двох або більше амінокислотних залишків, з’єднаних пептидними зв’язками. Пептиди, які складаються більше ніж з 10 амінокислотних залишків називаються, поліпептидами. Утворений дипептид може взаємодіяти з наступною амінокислотою, утворюючи трипептид, тетрапептид і т.д. Необхідно відмітити, що трипептид містить три залишки амінокислот, але не три пептидних зв’язки:
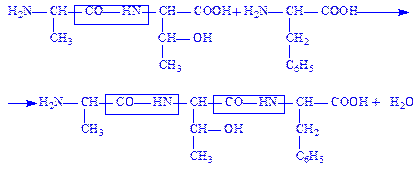
Структуру пептиду прийнято зображати так, щоб N-кінцевий залишок (містить вільну NH2–групу) розміщувався зліва, а С-кінцевий залишок (з вільною карбоксильною групою) – справа. Такий пептид має тільки одну вільну a-аміногрупу і тільки одну a-карбоксильну групу. Це твердження справедливе для всіх поліпептидів, які утворені тільки амінокислотними залишками, з’єднаними одна з одною пептидними зв’язками. В деяких пептидах кінцева аміногрупа або кінцева карбоксильна група модифікована (прикладом може бути ацильне похідне аміногрупи або амід карбоксильної групи), і таким чином, не являється вільною.
Лінійна послідовність амінокислотних залишків в поліпептидному ланцюзі називається первинною структурою пептиду. Для того, щоб встановити первинну структуру пептиду необхідно встановити число, хімічну структуру і порядок розміщення всіх амінокислотних залишків, які входять до його складу.
Заміна всього лише однієї амінокислоти на іншу в лінійній послідовності із 100 і більше залишків може призвести до зниження або до повної втрати біологічної активності пептиду, а це сприятиме розвиткові серйозних наслідків (наприклад, виникнення серповидної анемії).
Поліпептиди (білки) можуть містити 100 і більше амінокислотних залишків, тому традиційні структурні формули виявляються завеликими для позначення молекули пептиду. В даному випадку користуються “хімічним скорописом”, використовуючи трьохбуквенні або однобуквені позначення амінокислотних залишків (табл. 1). При найменуванні пептиду, його розглядають як похідне С-кінцевого амінокислотного залишку.
Водневий зв’язок досить поширений в хімічних сполуках. Він утворюється між ковалентно зв’язаним атомом водню, який має частковий позитивний заряд, та іншими ковалентно зв’язаними атомами, що мають негативний заряд. У молекулах білка водневий зв’язок найчастіше утворюється при взаємодії атома водню імінної групи залишку однієї амінокислоти з атомами кисню карбонільної групи залишку іншої амінокислоти:
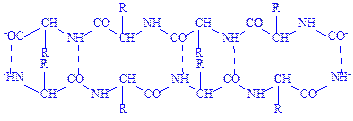
Водневий зв’язок у молекулі білка може бути внутрішньоланцюговим (з’єднує окремі витки однієї спіралі) і міжланцюговим (з’єднує різні поліпептидні ланцюги). У нативних білках цей тип зв’язку може утворюватися не лише між воднем і киснем пептидних груп, а й між іншими функціональними групами поліпептидного ланцюга.
Водневий зв’язок, на відміну від інших зв’язків, досить слабкий. Енергія цього зв’язку дорівнює 6 кДж, тоді як, наприклад, енергія зв’язку між атомами вуглецю становить близько 250 кДж. Тому водневий зв’язок легко утворюється і легко руйнується при звичайних умовах. Він відіграє важливу роль в утворенні вторинної, третинної і четвертинної структур білка.
Дисульфідний зв’язок. Відомо, що до складу переважної більшості білків входять залишки амінокислоти цистеїну. Цей досить міцний ковалентний зв’язок утворюється внаслідок відщеплення атомів водню від сульфгідрильних груп двох амінокислотних залишків цистеїну. Дисульфідний зв’язок, як і водневий, може бути внутрішньо ланцюговим (а) і міжланцюговим (б).
Схему утворення дисульфідного зв’язку можна показати так:
а)
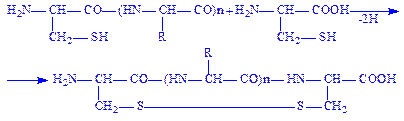
б)
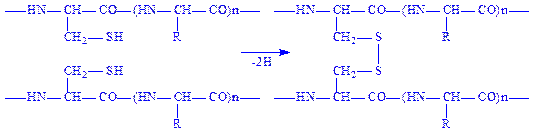
Дисульфідні зв’язки мають важливе значення для формування третинної структури білків. Руйнування цих зв’язків призводить до дестабілізації даного рівня структури і втрати білком його біологічної активності.
Іонний зв’язок утворюється при наявності у поліпептидних ланцюгах молекул білків залишків моноамінодикарбоновнх і діаміномонокарбонових кислот. Вільні карбоксильні та амінні групи цих залишків амінокислот перебувають переважно в іонізованому стані, в результаті чого між ними виникає електростатична взаємодія:
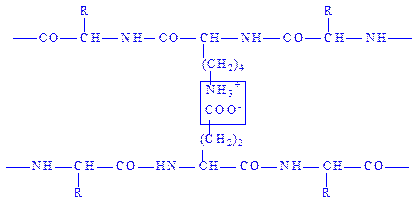
Іонний зв’язок відіграє важливу роль при утворенні третинної і, можливо, четвертинної структур білків.
Гідрофобний зв’язок утворюється внаслідок міжмолекулярної взаємодії (сил Ван-дер-Ваальса) між гідрофобними (неполярними) радикалами таких амінокислот, як аланін, валін, лейцин, ізолейцин, фенілаланін тощо:
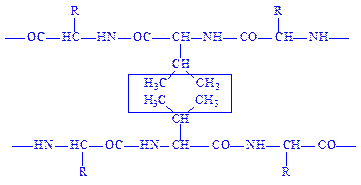
Гідрофобний зв’язок має важливе значення для стабілізації третинної і четвертинної структур білків.
Структури білків. Основополагаючим моментом в нашому розумінні принципів організації білків послужило виявлення того факту, що поліпептидні ланцюги можуть знаходитися у високо впорядкованій конформації, яка стабілізується водневими зв’язками між пептидними групами. Внаслідок існування таких конформацій розрізняють рівні структурної організації білка (рис. 2).
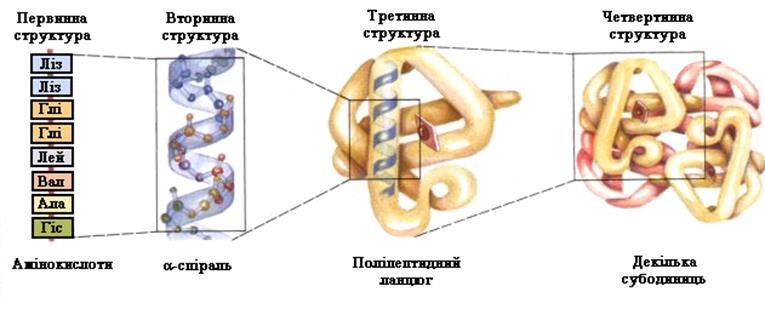
Рис. 2. Структури білкової молекули
Первинна структура білків – це послідовність амінокислот, зв’язаних пептидним зв’язком–СО–NH–. Унікальність первинної будови зумовлює функції та властивості білків, у тому числі імунні. Первинна будова білка кодується генетичним кодом у ДНК. Всі атоми пептидної групи компланарні, тобто знаходяться у одній площині, тому що мають делокалізовані p-електрони; О та Н розташовані у транс-положенні. Пептидний зв’язок існуючи на 90% в кетонній (транс-положення) та на 10% у енольній формах (таутомерія), здатен утворювати по два водневі зв’язки з полярними групами та з іншими пептидними зв’язками (зв’язки, що утворені проліном утворюють тільки один водневий зв’язок). Первинна структура білків є основою для самоорганізації цих молекул у просторі структури (рис. 3).
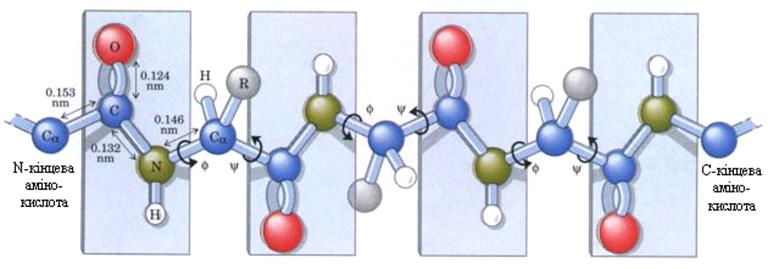
Рис. 3. Первинна структура білкової молекули
При вивченні первинної структури використовують як хімічні агенти, так і ферменти.
Протеолітичні ферменти. Це ферменти, що каталізують гідролітичний розрив пептидних зв'язків. Вони діляться на дві групи:
1) екзопептидази, які відщеплюють кінцеву амінокислоту, – амінопептидаза відділяє N-кінцеву амінокислоту, карбоксипептидаза відщеплює С-кінцеву амінокислоту;
2) ендопептидази, які діють на зв'язки між некінцевими амінокислотами. Ендопептидази володіють специфічністю відносно амінокислот, зв'язки яких гідролітично розриваються (з приєднанням елементів молекули води), – трипсин розриває молекулу на рівні карбоксилів лізину і аргініну; хімотрипсин розриває пептидні зв'язки тільки по карбоксилах ароматичних амінокислот: фенілаланіна, тирозина і триптофана.
Пепсин також є ендопептидазою, але його специфічність не цілком вивчена.
Визначення амінокислотного складу. Починають з повного гідролізу білка, тобто з розриву всіх пептидних зв'язків з приєднанням води і звільненням амінокислот. Гідроліз здійснюють дією 6N HCl при 110°C протягом 24 або 48 ч в запаяній під вакуумом ампулі (більш короткочасний гідроліз може виявитися неповним, при більш тривалому виникає небезпека руйнування невеликої частини амінокислот). Необхідно провести ще і лужний гідроліз невеликої кількості білка обробкою 2 – 4N NaOH для отримання триптофана, який при кислотному гідролізі руйнується. Отриману таким чином суміш амінокислот фракціонують і аналізують хроматографією на катіонообмінній смолі по описаному вище методу Мура і Стейна.
Визначення N-кінцевої амінокислоти. Для цієї мети можна використовувати різні методи (див. вище).
– метод Сенгера з динітрофторбензолом, ДНФБ.
– метод Едмана з використанням фенілізотіоціаната.
– дія амінопептидази.
Визначення С-кінцевої амінокислоти. Мається на увазі амінокислота, яка несе вільну COOH-групу і стоїть на кінці поліпептидного ланцюга. Її визначення досить важке (див. вище).
Визначення амінокислотної послідовності. Білок ділять на декілька зразків, які потім обробляють окремо. Один зразок, наприклад, гідролізують соляною кислотою на холоді, інший обробляють трипсином, третій хімотрипсином. Для кожного зразка одержують суміш пептидів різної довжини. Пептиди фракціонують методами електрофореза або хроматографії на іонообмінних смолах. Ці смоли схожі на ті, за допомогою яких фракціонують амінокислоти, але сітка полімеру в них менш тісна, що забезпечує проникання більш великих молекул всередину смоли і їх затримання.
Для кожного з поліпептидних фрагментів проводять обробку такого ж характеру, як для цілого білка, і якщо в результаті утворюються не дуже довгі пептиди, то вдається розшифрувати первинну структуру кожного з них, використовуючи, зокрема, метод Едмана. Потім встановлюють порядок взаємного розташування пептидів в молекулі білка. Цього досягають шляхом зіставлення пептидів, отриманих різними способами гідролізу, в яких амінокислотні послідовності частково перекриваються.
Метод „відбитків пальців” (фінгерпринт). Іноді виникає необхідність порівняти два близьких за своєю природою білка. Структура одного з них відома, і бажано визначити будову другого білка, не проводячи заново повного визначення його структури. В цьому випадку прагнуть вичленувати ті ділянки білків, які неідентичні. Для цього обидва білки спочатку піддають протеолізу, наприклад трипсином, а потім наносять в кут листа паперу гідролізат і проводять хроматографічне розділення в одному напрямі і електрофорез в перпендикулярному напрямі. Папір висушують і фарбують пептиди, що розділилися на площі листа. При цьому виявляють, що більшість плям розташована однаково, тобто, відповідає однаковим пептидам, і лише один або декілька пептидів – відмінні. Ці пептиди походять з тих частин молекул двох білків, які не ідентичні. Їх можна проаналізувати.
Цим методом Інгрему вдалося зіставити два гемоглобіни (білки, утворені поліпептидними ланцюгами двох типів, кожен з яких містить більше 140 амінокислот). Один з гемоглобінів був нормальним, а другий – виділений з крові хворого дрепаноцитозом (захворювання крові, відоме також під назвою серповидноклітинної анемії). Відмінність між двома гемоглобінами виявилася тільки в одному пептиді і була обумовлена заміною лише однієї амінокислоти.
Вторинна структура білка – це просторове розміщення первинної будови у вигляді a-спіралі або b-складчатої будови. Стабілізується вторинна структура численними водневими зв’язками між різними пептидними групами =C=O...H–N=. Спіральна конфігурація a-спіралі має гвинтову симетрію, водневі зв’язки утворюються між пептидними групами першим та четвертим амінокислотних залишків, причому бокові радикали не приймають участь у утворенні a-спіралі. Звичайно a-спіраль є у більшості білків, або ж як фрагмент молекули білка. b-складчаста структура зустрічається у структурних білків, частіше у білку містяться обидві структури.

