2. Производные гемоглобина. Аномальные гемоглобины. Серповидно-клеточная анемия и талассемия
Мутантные (М) гемоглобины человека
Мутации генов, кодирующих α- и β-цепи, могут существенным образом сказываться на их биологической функции. Известно несколько сот мутантных гемоглобинов человека (в большинстве случаев функционально активных), и о некоторых из них, отличающихся сильным изменением биологических функций, речь пойдет ниже. Патологическое состояние, при котором мутация вызывает изменение биологической функции гемоглобина, называют гемоглобинопатией.
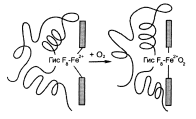
В семействе гемоглобинов М остатки проксимального или дистального гистидина в α- или β-субъединицах заменены на остатки тирозина. Атом железа в составе гема находится в этом случае в Fe3+-состоянии, что обусловлено образованием прочного ионного комплекса с фенолятным анионом тирозина. Результатом такой аномалии является метгемоглобинемия, поскольку ферри-гем не способен связывать О2. В α-цепи гемоглобина М R-Т-равновесие сдвинуто в сторону образования Т-формы. Сродство к кислороду низкое, эффект Бора отсутствует.
В β-цепях гемоглобинов М может происходить переход между R- и Т-состояниями и, следовательно, наблюдается эффект Бора. Мутации, приводящие к преимущественному образованию R-формы (в качестве примера можно привести гемоглобин Чезапик), отличаются тем, что соответствующие гемоглобины обладают повышенным сродством к кислороду. Подобные гемоглобины не способны поставлять достаточное количество кислорода периферическим тканям. Возникает тканевая гипоксия, ведущая к развитию полицитемии (повышению концентрации эритроцитов).
Талассемии
Другая важная группа нарушений, связанных с аномалиями гемоглобина – талассемии. Для них характерна пониженная скорость синтеза α-цепей гемоглобина (α-талассемия) или β-цепей (β-талассемия). Это приводит к анемии, которая может принимать очень тяжелую форму. В последние годы достигнут ощутимый прогресс в выяснении молекулярных механизмов, ответственных за развитие талассемии.
Гемоглобин при серповидноклеточнон анемии
В гемоглобине S остаток Glu A2(6)β замещен на Val. Остаток А2 (Glu или Val) располагается на поверхности молекулы гемоглобина и контактирует с водой, и замещение полярного остатка Glu на неполярный Val приводит к появлению на поверхности β-субъединицы «липкого участка». Этот липкий участок присутствует как в оксигенированном, так и в дезоксигенированном гемоглобине S (в гемоглобине А он отсутствует). На поверхности дезоксигенированного гемоглобина существует комплементарный участок, способный прочно связываться с липким участком β-субъединицы, тогда как в оксигенированном гемоглобине этот участок маскируется другими группами. Когда гемоглобин S переходит в дезоксигенированное состояние, его липкий участок связывается с комплементарным участком на другой молекуле дезоксигенированного гемоглобина.
Происходит полимеризация дезоксигемоглобина S и его осаждение в виде длинных волокон. Волокна дезоксигемоглобина S механически деформируют эритроцит, придавая ему серповидную форму, что приводит к лизису клеток и множеству вторичных клинических проявлений.
Таким образом, если бы можно было поддерживать гемоглобин S в оксигенированном состоянии или по крайней мере свести к минимуму концентрацию дезоксигенированного гемоглобина S, то нам удалось бы предотвратить полимеризацию дезоксигенированного гемоглобина S и образование «серповидных» клеток. Ясно, что полимеризации подвержена Т-форма гемоглобина S. Интересно отметить (хотя в практическом плане это мало существенно), что ферри-ион метгемоглобина А остается в плоскости порфиринового кольца и тем самым стабилизирует R-форму гемоглобина. То же относится и к гемоглобину при серповидноклеточной анемии: гемоглобин S в ферри-состоянии (метгемоглобин S) не подвержен полимеризации, поскольку он стабилизирован в R-форме.
В дезоксигемоглобине А тоже имеется рецепторный участок, способный взаимодействовать с липким участком оксигенированного или дезоксигенированного гемоглобина S, но присоединения «липкого» гемоглобина S к дезоксигемоглобину А недостаточно для образования полимера, поскольку сам дезоксигемоглобин А липкого участка не содержит и не может связать следующую молекулу гемоглобина. Следовательно, связывание дезоксигемоглобина А с R- или Т-формой гемоглобина S прерывает полимеризацию.
В результате полимеризации дезоксигемоглобина S образуются спиральные фибриллярные структуры. При этом каждая молекула гемоглобина контактирует с четырьмя соседними молекулами. Образование подобных трубчатых волокон ответственно за механические нарушения в содержащем их эритроците: он приобретает серповидную форму, становится подверженным лизису в момент прохождения им щелей в синусоидах селезенки.
3. Конформационные изменения и кооперативные взаимодействия субъединиц гемоглобина. Эффект Бора. Роль 2,3–бисфосфоглицерата
Кооперативные изменения конформации гемоглобина
В дезоксигемоглобине благодаря ковалентной связи с белковой частью атом Fe2+ выступает из плоскости гема в направлении Гис F8. Присоединение О2 к атому Fe2+ одного протомера вызывает его перемещение в плоскость гема, за ним перемещаются остаток Гис F8 и полипептидная цепь, в состав которой он входит. Т.к. протомер связан с остальными протомерами, а белки обладают конформационной лабильностью, происходит изменение конформации всего белка.
Конформационные изменения, произошедшие в других протомерах, облегчают присоединение следующей молекулы О2, что вызывает новые конформациолнные изменения в белке и ускорение связывания следующей молекулы О2. Четвёртая молекула О2 присоединяется к гемоглобину в 300 раз легче, чем первая молекула. Аналогичным образом в тканях диссоциация каждой молекулы О2 изменяет конформацию всех протомеров и облегчает отщепление последующих молекул О2.
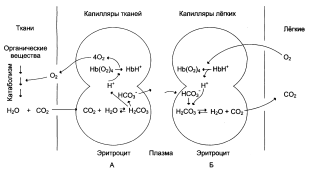
Молекулярная основа эффекта Бора
Протоны, ответственные за эффект Бора, высвобождаются в результате разрушения солевых мостиков, которым сопровождается связывание кислорода с Т-структурой; они отсоединяются от атомов азота остатков гистидина (146) в β-цепях. Эти протоны сдвигают равновесие в сторону образования угольной кислоты, которая расщепляется карбоангидразой с образованием СО2. Наоборот, при высвобождении кислорода вновь формируется Т-структура с присущими ей солевыми мостиками, при образовании которых происходит присоединение протонов к остаткам гистидина в β-цепях. Таким образом, в периферических тканях протоны благоприятствуют образованию солевых мостиков путем протонирования (по атому азота) концевых остатков гистидина в C-субъединицах. Образование солевых мостиков форсирует освобождение кислорода из оксигенированной R-формы гемоглобина. Итак, повышение концентрации протонов способствует освобождению кислорода, а повышение концентрации кислорода стимулирует высвобождение протонов. Первый из этих эффектов проявляется в сдвиге кривой диссоциации кислорода вправо при повышении концентрации ионов водорода (протонов).
Регуляция 2,3-бисфосфоглицератом сродства гемоглобина к О2
Недостаток кислорода в периферических тканях приводит к накоплению 2,3-бисфосфоглицерата (дифосфоглицерата, ДФГ). Это соединение образуется из 1,3-бисфосфоглицерата, промежуточного продукта гликолиза. Тетрамер гемоглобина связывает одну молекулу ДФГ, которая размещается в центральной полости, выстланной остатками всех четырех субъединиц. Объем этой полости достаточен для размещения ДФГ только в том случае, когда молекула гемоглобина находится в Т-форме и образуется достаточно широкий просвет между Н-спиралями β-цепей. Связывание ДФГ осуществляется путем образования солевых мостиков между атомами кислорода ДФГ и группами, принадлежащими обеим C-цепям. Таким образом, ДФГ стабилизирует дезоксигенированную Т-форму гемоглобина, образуя поперечные связи между β-цепями – дополнительные солевые мостики, которые должны быть разрушены при переходе гемоглобина из Т- в R-форму. 2,3-бисфосфоглицерат является аллостерическим регулятором сродства гемоглобина к кислороду.
В крови, консервированной в цитрат-декстрозной среде, за 10 дней концентрация 2,3-бисфосфоглицерата снижается с 4,5 до 0,5 мМ. Гемоглобин такой крови имеет очень высокое сродство к кислороду. Если кровь со сниженной концентрацией БФГ переливать тяжелобольным, возникает опасность развития гипоксии тканей.
Изменение положения Fe2+ и белковой части гемоглобина при присоединении О2
Кооперативные изменения конформации протомеров гемоглобина при присоединении О2

 Почему я выбрал профессую экономиста
Почему я выбрал профессую экономиста
 Почему одни успешнее, чем другие
Почему одни успешнее, чем другие
 Периферийные устройства ЭВМ
Периферийные устройства ЭВМ
 Нейроглия (или проще глия, глиальные клетки)
Нейроглия (или проще глия, глиальные клетки)

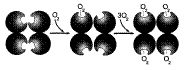
 | 4398 -
| 4398 - 