Тема 1. Введение. Строение и функции белков и аминокислот
Лекция 3. Гемоглобин и миоглобин. Протеомика. Переваривание белков.
Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного и количественного состава в крови. Гемоглобин АIС и его клиническое значение.
Производные гемоглобина. Аномальные гемоглобины. Серповидно-клеточная анемия и талассемия.
Конформационные изменения и кооперативные взаимодействия субъединиц гемоглобина. Эффект Бора. Роль 2,3–бисфосфоглицерата.
Роль протеомики в оценке патологических состояний. Понятие об азотистом балансе. Переваривание и всасывание пищевых белков. Протеиназы желудочно-кишечного тракта: пепсин, гастриксин, ренин, трипсин, химотрипсин, экзопептидазы. Гастроинтестинальные гормоны, их роль.
Бактериальное расщепление невсосавшихся в кишечнике аминокислот.
| Краткое содержание | Конспект | ||||||||
| 1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного и количественного состава в крови. Гемоглобин АIС и его клиническое значение | |||||||||
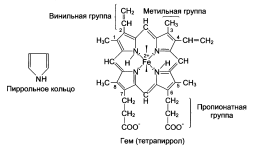
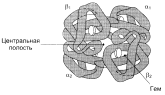
| К группе гемопротеинов относятся гемоглобин и его производные, миоглобин, хлорофиллсодержащие белки и ферменты (вся цитохромная система, каталаза и пероксидаза). Все они содержат в качестве небелкового компонента структурно сходные железо- (или магний) порфирины, но различные по составу и структуре белки, обеспечивая тем самым разнообразие их биологических функций. Гемоглобин в качестве белкового компонента содержит глобин, а небелкового – гем. Видовые различия гемоглобина обусловлены глобином, в то время как гем одинаков у всех видов гемоглобина. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Последний состоит из четырех замещенных пирролов, соединенных между собой метиновыми мостиками (—СН=). Незамещенный порфирин называется порфином. В молекуле гема порфин представлен в виде протопорфирина IX, содержащего четыре метильные группы (—СН3), две винильные группы (—СН=СН2) и два остатка пропионовой кислоты. Протопорфирин, присоединяя железо, превращается в гем. Гем в виде гем-порфирина является простетической группой не только гемоглобина и его производных, но и миоглобина, каталазы, пероксидазы и цитохромов b, с и c 1. Атом железа расположен в центре гема-пигмента, придающего крови характерный красный цвет. Каждая из 4 молекул гема «обернута» одной полипептидной цепью. В молекуле гемоглобина взрослого человека HbА (от англ. adult – взрослый) содержатся четыре полипептидные цепи, которые вместе составляют белковую часть молекулы – глобин. Две из них, называемые α-цепями, имеют одинаковую первичную структуру (по 141 аминокислотному остатку). Две другие, обозначаемые β-цепями, также идентично построены (содержат по 146 аминокислотных остатков). Таким образом, вся молекула белковой части гемоглобина состоит из 574 аминокислот. Во многих положениях α- и β-цепи содержат разные аминокислотные последовательности, хотя и имеют почти одинаковые пространственные структуры. Миоглобин –содержится в красных мышцах и участвует в запасании кислорода. В условиях интенсивной мышечной работы, когда парциальное давление кислорода в ткани падает, О2 освобождается из комплекса с миогобином и используется в митохондриях клеток для получения необходимой для работы мышц энергии. Гемоглобины – структурно-родственные белки, находящиеся в эритроцитах позвоночных. Онивыполняют две важные биологические функции: 1) переносят О2 из легких к периферическим тканям; 2) переносят СО2 и протоны от периферических тканей к дыхательным органам для последующего выведения из организма. Гемоглобин связывает четыре молекулы кислорода на тетрамер (по одной на гем в каждой субъединице); особенно важным отличием его от миоглобина является характерная кривая насыщения кислородом, которая имеет сигмоидную форму. Таким образом, способность гемоглобина связывать О2 зависит от того, содержатся ли в данном тетрамере другие молекулы О2. Если да, то последующие молекулы О2 присоединяются легче. Следовательно, для гемоглобина характерна кинетика кооперативного связывания, благодаря которой он связывает максимальное количество О2 в легких и отдает максимальное количество О2 при тех Ро, которые имеют место в периферических тканях. Гемоглобин не только переносит кислород от легких к периферическим тканям, но и ускоряет транспорт СО2 от тканей к легким. Гемоглобин связывает СО2 сразу после высвобождения кислорода; примерно 15% СО2, присутствующего в крови, переносится молекулами гемоглобина. Находящаяся в эритроцитах карбоангидраза катализирует превращение поступающего из тканей СО2 в угольную кислоту. Угольная кислота быстро диссоциирует на бикарбонат-ион и протон, причем равновесие сдвинуто в сторону диссоциации. Для предотвращения опасного повышения кислотности крови должна существовать буферная система, способная поглощать избыток протонов. Гемоглобин связывает два протона на каждые четыре освободившиеся молекулы кислорода и определяет буферную емкость крови. В легких идет обратный процесс: присоединение кислорода к дезоксигемоглобину сопровождается высвобождением протонов, которые связываются с бикарбонат-ионами, переводя их в угольную кислоту. Далее эффективно действующая карбоангидраза катализирует превращение угольной кислоты в углекислый газ, выдыхаемый из легких. Таким образом, связывание кислорода тесно сопряжено с выдыханием СО2. Это обратимое явление известно как эффект Бора. Эффект Бора является свойством тетрамерного гемоглобина и определяется гем-гемовым взаимодействием, лежащим в основе кооперативных эффектов. У миоглобина эффект Бора не обнаруживается. Наиболее распространенные гемоглобины имеют следующую тетрамерную структуру: · НЬА (нормальный гемоглобин взрослого человека) – α2β2. Это основной гемоглобин взрослого организма, составляет около 98% от общего количества гемоглобина; · HbA2 (минорный гемоглобин взрослого человека) – α2δ2 Он находится в организме взрослого человека в меньшей концентрации, на его долю приходится около 2% общего гемоглобина; · Гемоглобин АIc – гемоглобин А, модифицированный ковалентным присоединением к нему глюкозы (гликозилированный гемоглобин); · Эмбриональный гемоглобин – синтезируется в эмбриональном желточном мешке через несколько недель после оплодотворения – ξ2ε2. Через 2 недели после формирования печени плода в ней начинает синтезироваться гемоглобин F; · HbF (фетальный гемоглобин) – α2γ2; · HbS (гемоглобин при серповидно-клеточной анемии) – α2S2. Четвертичная структура наделяет гемоглобин дополнительными важными особенностями (отсутствующими у миоглобина), которые способствуют выполнению гемоглобином его уникальной биологической функции и обеспечивают возможность строгой регуляции его свойств. |
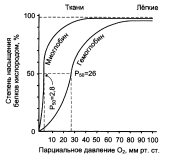
Из графика видно, что гемоглобин имеет значительно более низкое сродство к О2. Полунасыщение гемоглобина О2 наступает при более высоком давлении кислорода (около 26 мм рт.ст.). Парциальное давление кислорода в артериальной крови составляет около 100 мм рт. ст., в венозной крови около 40 мм рт. ст., в капиллярах кровеносных сосудов активной мышцы – около 20 мм рт. ст.; минимальное давление, необходимое для функционирования ферментов цитохромной системы, равно ~5 мм рт. ст. Из рисунка видно, что ассоциация цепей с образованием тетрамерной структуры приводит к существенному повышению эффективности снабжения тканей кислородом по сравнению с мономерными белками. Ход кривой насыщения гемоглобина кислородом или диссоциации оксигемоглобина зависит от ряда факторов: 1. Сродство гемоглобина к кислороду в первую очередь связано с рН. Чем ниже рН, тем меньше способность гемоглобина связывать кислород и тем выше Р50. В тканевых капиллярах рН ниже (поступает большое количество СО2), в связи с чем гемоглобин легко отдает кислород. В легких СО2 выделяется, рН повышается и гемоглобин активно присоединяет кислород. 2. Способность гемоглобина связывать кислород зависит также от температуры. Чем выше температура (в тканях температура выше, чем в легких), тем меньше сродство гемоглобина к кислороду. Напротив, снижение температуры вызывает обратные явления. 3. Количество гемоглобина в крови, а также в какой-то мере его способность связывать кислород (характер кривой диссоциации оксигемоглобина) несколько меняются с возрастом. Например, у новорожденных содержание гемоглобина доходит до 20-21% (вместо обычных для взрослого 13-16%). У человека имеется несколько гемоглобинов, которые образуются в различном количестве в разные стадии онтогенеза и различаются по своему сродству к кислороду.
Содержание гемоглобина А в крови взрослого здорового человека: · женщины - ……… · мужчины - ……... |
 А Б
А Б