




Локальний нервовий центр забезпечує підтримання констант газового складу крові в стані спокою.
Локальний нервовий центр складається з
- дорсального й вентрального ядер довгастого мозку (медулярний відділ),
- пневмотаксичного (міст)
- апнейстичного відділів (міст).
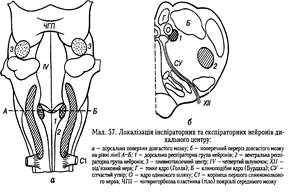
Частина, яка розташована у ділянці довгастого мозку називається медулярною. Вона має два скупчення ядер:
- дорсальне,
- вентральне.
Ці скупчення нейронів проявляють свою активність у момент здійснення вдиху або видиху. Якщо збудження нейрона збігається із вдихом, то такий нейрон належить до інспіраторних, якщо із видихом - до експіраторних. Нейрони цих ядер широко контактують з ретикулярною формацією мозкового стовбура.
Дорсальне ядро містить нейрони, що збуджуються під час інспірації(вдиху). У ньому можна виділити два основні типи нейронів:
- Іа - нейрони, які збуджуються тільки під час вдиху;
- Ір - нейрони, що збуджуються і одночасно з Іа і під час паузи останніх.
Іа-нейрони є типовими інспіраторними нейронами. Нервові імпульси від них передаються на мотонейрони діафрагми, що розташовані в спинному мозку (3-й і 4-й шийні сегменти). Одночасно із цим збудження Іа-нейронів передається до ір-нейронів. Однак вони не передають свої імпульси до мотонейронів діафрагми і їх збудження гальмує активність Іа-нейронів.
Верхня частина вентрального ядра містить більше інспіраторних нейронів, а нижня - експіраторних. Ураховуючи це, вентральне ядро можна поділити на два відділи:
- інспіраторний,
- експіраторний.
Велика частина нервових волокон від цих ядер йде до грудних сегментів спинного мозку, до мотонейронів міжребрових м'язів і м'язів живота (відповідно до м'язів, що забезпечують вдих чи видих).
Лише 20-25% нервових волокон розгалужується у ділянці діафрагмальних ядер.
Крім названих бульбарних центрів у передній частині моста виявлено ще одне ядро, яке бере участь у регулюванні дихання. Це - пневмотаксичний центр, який, впливаючи на бульбарний відділ дихального центру, забезпечує регулювання зміни та тривалість вдиху і видиху.
Деякі автори вважають, що в нижніх відділах моста розташований ще й апнейстичний центр, імпульсація якого збуджує ір-нейрони дорсального ядра.
Пневмотаксичний центр.
У верхній частині мосту (варолієвого) розміщені скупчення дихальних нейронів, які функціонально відрізняються від дихальних нейронів довгастого мозку і утворюють пневмотаксичний центр.
Вони не жорстко пов'язані з дихальним ритмом, можуть змінювати свою функцію: з інспіраторних перетворюватись на експіраторні і навпаки.
Велика кількість нейронів активізується в перехідних фазах від вдиху до видиху чи від видиху до вдиху. Подразнення пневмотаксичного центру може спричинювати як вдих, так і видих.
Пневмотаксичний центр отримує імпульси від інспіраторного центру, а свої сигнали надсилає до обох — інспіраторного та експіраторного центрів. Вважають, що пневмотаксичний центр перемикає збудження з інспіраторних нейронів на експіраторні і модулює ритм дихання, пристосовуючи його до потреб організму.
Вище згадувалось, що зміна вдиху на видих відбувається за допомогою рефлексу Герінга —Бреєра (це рефлексвід блукаючих нервів, через які сигнали від механорецепторів легень надходять до дихального центру). Якщо цей рефлекс припинити перерізуванням обох блукаючих нервів, дихання не припиниться, але стане значно рідшим. Ритм дихання підтримуватиметься пневмотаксичним центром. Його зруйнування у ваготомованої тварини призведе до зупинки дихання у фазі вдиху з дуже рідкими судомними видихами.
Отже, перехід від вдиху до видиху забезпечується подвійним механізмом: рефлексами Герінга — Бреєра і пневмотаксичним центром.
Апнейстичний центр
(розміщений в нижній частині моста) активує інспіраторні нейрони дорзального ядра при гіпоксії з виникненням затяжного вдиху з наступним коротким видихом.
Він упереджує або припиняє розвиток гальмування вдиху. Крім того, збудження еферентними шляхами досягає мотонейронів дихальних м'язів (діафрагмального й зовнішніх косих міжреберних), спричиняючи їх скорочення.
Скорочення дихальних м'язів викликає акт першого вдиху, який настає через 20 с після народження. Ритмічне дихання встановлюється в межах 1,5 хв. Пневмотаксичний центр, гальмуючи дорзальні інспіраторні нейрони, зменшує тривалість вдиху і таким чином контролює наповнення легень повітрям та частоту дихання. Його активація збільшує частоту дихання до 40 за хвилину, гальмування - зменшує до кількох за хвилину.
Функціонування дихального центру
Головною особливістю дихального центру є періодичність його діяльності, внаслідок якої збудження нейронів періодично змінюється на їх гальмування.
Основна роль у цьому процесі належить нейронам дорсального ядра Вважають, що своєрідним "водібм ритму" є Іа-нейрони. Від них збудження надходить до трьох відділів ЦНС:
- до мотонейронів шийного відділу спинного мозку,
- до інспіраторних нейронів вентрального ядра,
- до ф-нейронів, розташованих тут же у дорсальному ядрі.
Функція ір-нейронів полягає у гальмуванні Іа-нейронів і припиненні вдиху. Однак для "вимикання" Іа-нейронів необхідно, щоб рівень збудження ір-нейронів досягав визначеної критичної межі.
Гуморальна регуляція дихання.
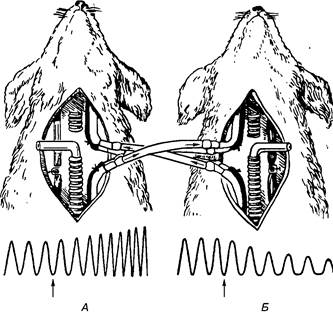
Наприкінці XIX ст. С. Фредерік здійснив дослід з перехресним кровообігом. Головний мозок, зокрема дихальний центр собаки А, отримує кров від собаки Б, а до голови останньої постачається кров від собаки А (мал. 59).
Якщо тепер спричинити асфіксію перетисканням трахеї у собаки Б, то дихання посилюватиметься у собаки А, а у першої тварини, навпаки, дихання буде гальмуватись. З цього досліду випливає висновок, що саме через кров дихальний центр отримує сигнали, які регулюють його функцію.
Таким сигналом є вуглекислий газ, зі збільшенням концентрації якого в крові підвищується тонус дихального центру, а зниженням, навпаки, гальмує його діяльність.
Зміна концентрації С02 в крові сприймається центральними хеморецепторами, розміщеними у нередньобічній ділянці довгастого мозку, а такий спосіб дії С02 безпосередньо через центральні хеморецептори дістав назву гуморальної регуляції дихання.
Пізнішими дослідженнями було встановлено, що С02 збуджує нейрони набагато слабше, ніж йони гідрогену. Проте Н+ майже не проникають крізь гематоенцефалічний бар'єр і не можуть перейти з крові до цереброспінальної рідини, що омиває мозок. Тому вважають, що у цереброспінальну рідину дифундує з крові С02 і, сполучаючись там з водою, утворює карбонатну (вугільну) кислоту, яка дисоціює з виділенням йонів гідрогену, які й діють безпосередньо на центральні хеморецептори довгастого мозку.
Активують вентиляцію гормональні фактори:
- адреналін,
- прогестерон.
- Особливе значення в стимуляції зовнішнього дихання мають деякі гуморальні компоненти крові. Так, підвищення рС02, зниження рН і р02 викликають посилення частоти й глибини дихальних рухів (гіперпное). Необхідно відмітити, що С02 є більш сильним подразником дихального центру, ніж рН і 02. Це підтверджується тим, що зменшення рН внаслідок накопичення в крові нелетких кислот (метаболічний ацидоз) і падіння рО2 при рС02 = 40 мм рт. ст. викликають менш суттєву стимуляцію зовнішнього дихання, ніж зменшення самого рС02. Підвищення рС02 до 60 мм рт. ст. викликає зростання хвилинного об'єму дихання, до 70 мм рт. ст. - зменшує вентиляцію.

