Синтез белка начинается в цитозоле на рибосомах. Рибосомные субъединицы покидают ядро в виде комплексов 40S и 60S. Эти субъединицы не образуют функционирующую рибосому до тех пор, пока не присоединятся к мРНК. Малая субъединица (40S) связывается с 5' участком мРНК вместе с другими компонентами комплексов трансляции белка. Затем присоединяется 60S субъединица и формируется зрелая 80S рибосома. Трансляция с мРНК начинается сразу после образования функционирующей рибосомы. В 60S субъединице есть канал, достаточно длинный для того, чтобы содержать полипептид, состоящий приблизительно из 30 аминокислотных остатков. Удлиняющийся полипептид проходит в этот канал.
Когда сформированная рибосома перемещается по нити мРНК, захватывая по 6 нуклеотидов, образуется второй, а затем третий комплексы трансляции, которые считывают информацию, заключенную в мРНК. Этот процесс продолжается до тех пор, пока рибосомные комплексы не «прочитают» всю мРНК.
Обычная мРНК связывается с 10-12 рибосомами, формируя таким образом полирибосомный комплекс, или полисому. Каждая рибосома, достигшая трансляционного конца, терминирующего кодона, покидает мРНК. Затем рибосомные субъединицы отделяются друг от друга и поступают в цитозольный пул, из которого впоследствии могут быть образованы новые рибосомы. Эти рибосомы могут собираться на той же самой или другой молекуле мРНК. Рибосомные субъединицы могут использоваться многократно для трансляции любой нормальной мРНК.
Матричные РНК, подвергающиеся трансляции, делятся на две функциональные группы.
1. мРНК, кодирующие секреторные белки или интегральные белки мембраны. Эти молекулы направляются к поверхности эндоплазматического ретикулума.
2. мРНК, кодирующие белки, которые остаются в клетке. Эти синтезирующие комплексы остаются в цитозоле, не прикрепляясь к мембранам.
Необходимо отметить, что белки, синтезируемые на второй группе комплексов, составляют большую часть белков, синтезируемых в клетке. Мы уже видели, что некоторые из этих белков обладают адресными сигналами, составляющими часть их аминокислотной последовательности. Эти сигналы способствуют направленному движению белков в соответствующие клеточные органеллы.
Биосинтез мембранных и секреторных белков происходит в шероховатом ЭР. В этой клеточной органелле синтезируются все мембраны. Здесь также проходят начальные этапы посттрансляционного гликозилирования. Кроме того, укладка белка и сборка субъединиц также происходят в полости шероховатого ЭР.
Рибосомы — мелкие (15—30 нм) немембранные органоиды, встречающиеся в клетках всех живых организмов (рис. 2.11).
Количество рибосом в клетке может достигать нескольких миллионов. В рибосомах осуществляется синтез белка. Активно работающая рибосома имеет округлую или грибовидную форму и состоит из двух субъединиц: малой субъединицы, связывающей информационную РНК (мРНК), и большой субъединицы, катализирующей образование пептидных связей между аминокислотами. Всего в состав рибосом эукариотического типа входит около 80 молекул белка и 4 молекулы рибосомальной РНК (рРНК). Рибосомальная РНК синтезируется на дезоксирибонуклеиновой кислоте (ДНК) ядрышка, а рибосомные белки — в цитоплазме, откуда они транспортируются через ядерные поры в ядро. В ядрышке рибосомальные белки связываются с рРНК, образуя малые и большие субъединицы рибосом, которые затем по отдельности переносятся через ядерные поры из ядра в цитоплазму, где происходит формирование функционально зрелых рибосом.
Большая часть активно функционирующих рибосом (полисом) находится в цитоплазме в свободном состоянии, другая же часть их связана с мембранами эндоплазматической сети. Строение тех и других рибосом идентично, и они могут меняться местами в зависимости от функциональной активности клетки.
Рибосомы, находящиеся в цитоплазме в свободном состоянии, вырабатывают белки для нужд самой клетки. Здесь синтезируются белки цитозоля, ядерные белки, большинство митохондриальных белков, белки цитоскелета и пероксисом. Синтез всех перечисленных типов белков начинается и заканчивается в цитозоле, и затем oни посттрансляционно с помощью внутриклеточных белковых комплексов переносятся «по адресам». Такая адресность транспорта белков, синтезируемых свободными рибосомами, достигается благодаря тому, что все они имеют в своём составе специфические «сигнальные» последовательности аминокислот, в соответствии с которыми они распределяются в клетке.
Рибосомы, связанные с мембранами эндоплазматической сети, синтезируют мембранные белки клетки, ферменты лизосом, а также белки, которые предназначены на экспорт, в том числе белки внеклеточного матрикса.
Синтез рибосом в ядрышке
Ядрышко не окружено мембраной; это, скорее, скопление рибосомных частиц, образующееся в процессе сборки. Синтез рибосом — основной процесс, происходящий в ядре. Самые активные эукариотические клетки используют около 10 миллионов рибосом в течение одного клеточного цикла. Как часть структуры рибосомы, вокруг каждой рРНК субъединицы находится ряд высокоспециализированных белков. В состав малой субъединицы — 40S частицы — входят 30 уникальных белков, собранных вокруг молекулы 18S РНК. Большая субъединица — 60S частица — имеет 51 белок, связанный со своей главной молекулой 28S РНК. В комплекс большей субъединицы входит также 5,8S РНК.
Более подробно механизмы синтеза белка рассматриваются ниже в этой главе (рис. 3-4).

Ядерная оболочка
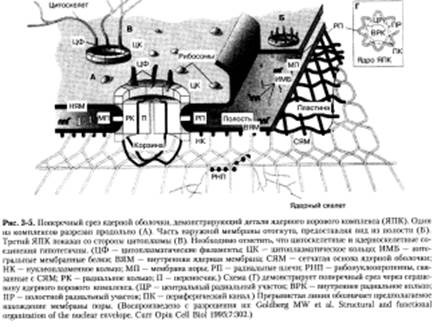
Ядерная оболочка — двойная мембранная структура, которая окружает хроматин и переходит в эндоплазматический ретикулум (ЭР). Внутренняя мембрана по составу белков отличается от наружной мембраны. Внутренний слой мембраны имеет волокнистую сеть белков, называемых ламинами, которые играют ключевую роль в поддержании структурной целостности мембраны. Наружная мембрана ядра переходит в мембрану ЭР и содержит белки, необходимые для связывания рибосом (рис. 3-5).