




Аммиак очень токсичное соединение, особенно для нервных клеток. При накоплении его возникает возбуждение нервной системы. Поэтому концентрация аммиака в организме должна сохраняться на низком уровне. Уровень аммиака в норме в крови не превышает 1-2 мг/л (это почти в 1000 раз меньше концентрации сахара в крови). На кроликах показано, что концентрация аммиака 50 мг/л является летальной. Таким образом, аммиак должен подвергаться связыванию в тканях с образованием нетоксичных соединений, легко выделяемых с мочой.
Аммиак образуется в ходе следующих процессов:
- дезаминирование аминокислот;
- дезаминирование биогенных аминов;
- дезаминирование пуриновых оснований (гуанина и аденина);
- дезаминирование глутамина и аспарагина;
- распад пиримидиновых оснований (урацила, тимина, цитозина). В тканях организма существуют следующие пути обезвреживания аммиака:
- образование мочевины (орнитиновый цикл);
- восстановительное аминирование кетокислот;
- образование амидов аминокислот - аспарагина и глутамина;
- образование аммонийных солей.
Главный путь обезвреживания аммиака - синтез мочевины, происходящий в печени.
В 1933 году Кребс и Гензелейт установили, что синтез мочевины представляет собой циклический процесс, в котором каталитическую роль играет орнитин. В последующие десятилетия были раскрыты все реакции этого процесса, и он получил название орнитинового цикла, или цикла Кребса-Гензелейта (рис. 39).

Рис. 39. Орнитиновый цикл мочевинобразования
Начальной реакцией этого цикла является синтез высокоэнергетического соединения - карбамоилфосфата под действием фермента - карбамоилфосфатсинтетазы:

Карбамоильная группа далее переносится на орнитин с образованием цитруллина; реакцию катализирует орнитин-карбамоилтрансфераза:

| Орнитин | Карбамоилфосфат | Цитруллин |
Затем цитруллин- реагирует с аспарагиновой кислотой, превращаясь в аргининянтарную кислоту при действии аргининосукцинатсинтетазы:

| Цитруллин | Аспартат | Аргининосукцинат |
Аргининосукцинат под действием аргининосукциназы распадается на аргинин и фумаровую кислоту:

| Аргининосукцинат | Аргинин | Фумарат |
Далее аргинин гидролизуется аргиназой с образованием мочевины и орнитина:

| Аргинин | Орнитин | Мочевина |
Мочевина поступает с током крови а почки и уносится из организма с мочой, а орнитин возвращается в цикл. Реакции орнитинового цикла до стадии образования цитруллина происходят в митохондриях, а последующие стадии - в цитозоле клеток. Если человек в сутки получает 80-100 г белков, то образуется 25-30 г мочевины.
Важным вспомогательным путем связывания аммиака является образование аспарагина и глутамина с участием аспарагинсинтетазы и глутаминсинтетазы соответственно:
аспартат + АТФ + NH3 = аспарагин + АМФ + Н4Р2О7;
глутамат + АТФ + NH3 = глутамин + АДФ + H3PО4.
Эти процессы активны в нервной и мышечной тканях, в почках, в печени.
Часть аммиака легко связывается с α-кетоглутаровой кислотой, т.е. протекает реакция, обратная окислительному дезаминированию глутаминовой кислоты:
α-кетоглутарат + NH3 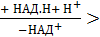 глутамат + Н2О
глутамат + Н2О
Но вклад этой реакции в обезвреживание аммиака невелик, так как необходимы значительные количества α-кетоглутарата.
Глутамин и в меньшей степени аспарагин считают как бы транспортной формой аммиака, так как, образуясь в тканях, они с кровью попадают в почки, где подвергаются гидролизу под действием специфических ферментов - глутаминазы и аспарагиназы:
аспарагин + Н2О = аспартат + NH3;
глутамин + H2O = глутамат + NH3.
Освободившийся в канальцах почек аммиак нейтрализует кислые продукты обмена при ацидозе, защищая тем самым организм от потери с мочой ионов Na+ и К*. При этом образуются соли аммония:
NH3 + Н+ + СI¯ = NH4CI,
которые выделяются с мочой.
Синтез аминокислот
В организме может синтезироваться любая из заменимых аминокислот. При этом исходными веществами при биосинтезе заменимых аминокислот служат метаболиты обмена углеводов.
Установлено, что в живых организмах путем реакций прямого восстановительного аминирования синтезируется три аминокислоты: аланин, аспартат и глутамат из пирувата, оксалоацетата и α-кетоглутарата соответственно:

| Пируват | Аланин |

| Оксалоацета | Аспартат |

| α-кетоглутарат | Глутамат |
Остальные аминокислоты, как правило, получаются в результате реакций трансаминирования вышеназванных аминокислот с соответствующими кетокислотами. Поэтому аланин, аспартат и глутамат часто называют первичными, а остальные аминокислоты - вторичными. Первичные аминокислоты могут синтезироваться также и в результате реакций трансаминирования:
| пируват + глутамин | 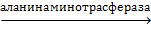

| аланин + α-кетоглутарат |
| Оксалоацета + глутамат |  
| Аспартат + α-кетоглутарат |
Глутамин и аспарагин с участием глутаминсинтетазы и аспарагинсинтетазы синтезируются по уравнениям:
аспартат + АТФ + NH3 = аспарагин + АМФ + Н4Р2О7
глутамат + АТФ + NH3 = глутамин + АДФ + Н3РО4
Аспарагин синтезируется также из аспартата и глутамина, который служит донором амидной группы:

| аспартат | глутамин | аспарагин | глутамат |
Пролин образуется из глутаминовой кислоты:

| Глутамат | γ-полуальдегид глутамата | Пролин |
Из 3-фосфоглицерата образуется серин, а из серина - глицин. Поэтому общим предшественником для серина и глицина можно считать 3-фосфоглицерат:

серии + ТГФК = глицин + N5, N10-метилен-ТГФК.
(ТГФК - тетрагидрофолиевая кислота)
Из рибозо-5-фосфата образуется гистидин. Первоначально рибозо-5-фосфат превращается в α-5-фосфорибозил-1-дифосфат (ФРДФ)(см. главу 12), который участвует в дальнейших стадиях биосинтеза и гистидина, и пуринов:
рибозо-5-фосфат → ФРДФ → гистидин.
Однако возможности биосинтеза гистидина таким способом ограничены, поэтому он является полузаменимой аминокислотой.
Из незаменимой аминокислоты фенилаланина образуется заменимая - тирозин. Это процесс катализируется фенилаланингидроксилазой, кофактором которой является дигидробиоптерин, а восстановителем – НАД.Н+Н+.
Фенилаланин + НАДФ.Н + Н+ + О2 = тирозин + НАДФ+ + Н2О
Поскольку фенилаланин - незаменимая кислота, то возможности эндогенногосинтеза тирозина ограничены. Метионин (незаменимая кислота) может превращаться в цистеин.
Орнитин в животных тканях превращается в аргинин. Однако эта возможность образования аргинина невелика, так как он сам служит субстратом для синтеза мочевины. Поэтому аргинин является полузаменимой аминокислотой.
Незаменимые аминокислоты, за исключением лизина и треонина, участвуют в реакциях трансаминирования, следовательно, при наличии соответствующих α-кетокислот они тоже могли бы синтезироваться в организме. Незаменимы собственно α-кетокислоты, соответствующие незаменимым аминокислотам. Единственным источником таких α-кетокислот служат незаменимые аминокислоты пищи. Поэтому трансаминирование незаменимых аминокислот служит только этапом их катаболизма, а не синтеза, в отличие от заменимых аминокислот, для которых трансаминирование может быть начальной стадией катаболизма или конечной стадией синтеза.

