




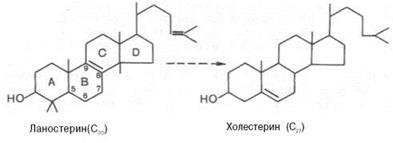
все атомы углерода холестерина происходят из ацетата. В синтезе холестерина можно выделить три основные стадии: I – превращение активного ацетата в мевалоновую кислоту, II – образование сквалена из мевалоновой кислоты, III – циклизация сквалена в холестерин.
ГМГ-КоА-редуктазная реакция – первая практически необратимая реакция в цепи биосинтеза холестерина. Она протекает со значительной потерей свободной энергии (около 33,6 кДж). Установлено, что данная реакция лимитирует скорость биосинтеза холестерина.
Начиная со сквалена, все промежуточные продукты биосинтеза холестерина (включая и холестерин) нерастворимы в водной среде. Поэтому они участвуют в конечных реакциях биосинтеза холестерина, будучи связанными со стеринпереносящими белками (СПБ). Это обеспечивает их растворимость в цитозоле клетки и протекание соответствующих реакций. Данный факт имеет важное значение и для вхождения холестерина в клеточные мембраны, окисления в желчные кислоты, превращения в стероидные гормоны. Как отмечалось, реакцией, регулирующей скорость биосинтеза холестерина в целом, является восстановление β-гидрокси-β-метилглутарил-КоА в мевалоновую кислоту, катализируемое ГМГ-КоА-редуктазой. Данный фермент испытывает регуляторное воздействие ряда факторов. В частности, скорость синтеза редуктазы в печени подвержена четким суточным колебаниям: максимум ее приходится на полночь, а минимум – на утренние часы. Активность ГМГ-редуктазы возрастает при введении инсулина и тире-оидных гормонов. Это приводит к усилению синтеза холестерина и повышению его уровня в крови. При голодании, тиреоидэктомии, введение глюкагона и глюкокорти-коидов, напротив, отмечается угнетение синтеза холестерина, что прежде всего связано со снижением активности ГМГ-КоА-редуктазы.
Желчные кислоты
ЖЕЛЧНЫЕ КИСЛОТЫ, монокарбоновые гидроксикислоты, относящиеся к классу стероидов. Почти все желчные кислоты - производные прир. холановой к-ты, синтезируемой из холестерина в печени.

Сначала синтезируются две первичные желчные кты – холевая и хенодезоксихолевая. Потом вторичные: дезоксихолевая и литохолевая. Функционируют они в виде конъюгантов с глицином или таурином. Конъюганты в печени образуются в две стадии:
R-COOH+ATP+KoASH = R – CO~SKoA+AMP+PP
R-CO~SKoA+H2N-CH2-COOH(глицин) = R-CO-NH-CH2-COOH+KoASH
Или
R-CO~SKoA+ H2N-CH2-CH2-SO3H(таурин) = R-CO-NH-CH2-CH2-SO3H+KoASH
Желчные кислоты эмульгируют жиры.
Стероидные гормоны
Общим предшественником стероидных гормонов является холестерин.
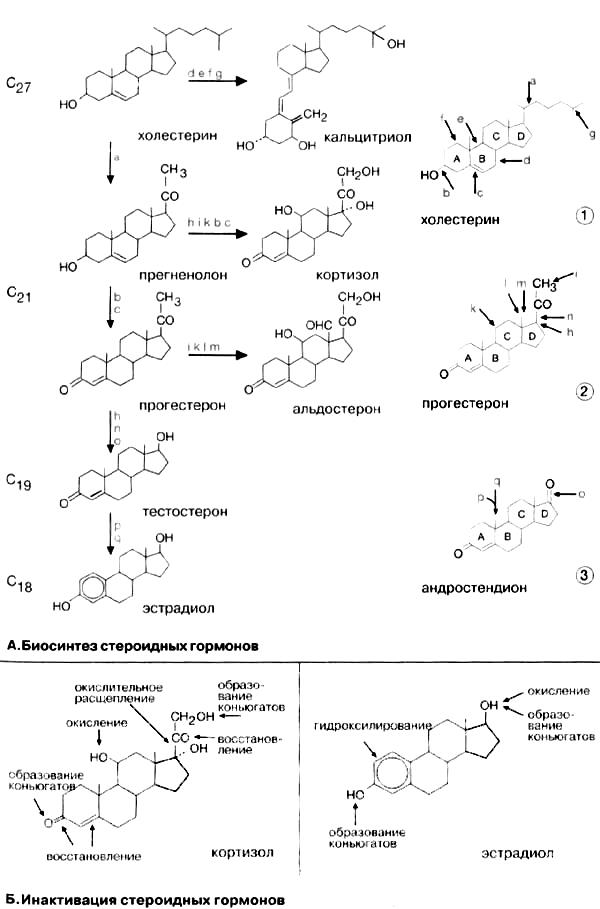
Путь биосинтеза. Биосинтез каждого гормона состоит из множества последовательных ферментативных реакций. В качестве примера рассмотрим биосинтез прогестерона (А) см. рис. 397). Биосинтез начинается с расщепления боковой цепи холестерина между С-20 и С-22 (а). Стероидное соединение с укороченной боковой цепью носит название прегненолон. Последующие стадии, окисление гидроксигруппы при С-3 (b) и сдвиг двойной связи от С-5 к С-4 (с) приводят к образованию прогестерона.
Приведенные на схеме стероиды объединены в подгруппы по числу углеродных атомов. Холестерин и кальцитриол являются С27-стероидами. Соединения с укороченной на 6 атомов углерода боковой цепью, прогестерон, кортизол и альдостерон, составляют группу С21-стероидов. В ходе биосинтеза тестостерон полностью утрачивает боковую цепь и поэтому его относят к С19-стероидам. При биосинтезе эстрадиола на стадии образования ароматического цикла теряется ангулярная метильная группа и, следовательно, эстрадиол является С18-стероидом.
В процессе биосинтеза кальцитриол подвергается фотохимической реакции раскрытия кольца В. Поэтому его относят к «секостероидам». Однако по своим биохимическим свойствам он является типичным стероидным гормоном.

