




система структурно и функционально связанных трансмембранных белков и переносчиков электронов. ЭТЦ позволяет запасти энергию, выделяющуюся в ходе окисления НАД•Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану.
У прокариот ЭТЦ локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики расположены по своему окислительно-восстановительному потенциалу, транспорт электрона на всём протяжении цепи протекает самопроизвольно.
Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.
-Комплекс I (НАДН дегидрогеназа) окисляет НАД-Н, отбирая у него два электрона и перенося их на растворимый в липидах убихинон, который внутри мембраны диффундирует к комплексу III. Вместе с этим, комплекс I перекачивает 4 протона из матрикса в межмембранное пространство митохондрии.
-Комплекс II (Сукцинат дегидрогеназа) не перекачивает протоны, но обеспечивает вход в цепь дополнительных электронов за счёт окисления сукцината.
-Комплекс III (Цитохром bc1 комплекс) переносит электроны с убихинола на два водорастворимых цитохрома с, расположенных на внутренней мембране митохондрии. Убихинол передаёт 2 электрона, а цитохромы за один цикл переносят по одному электрону.
При этом туда также переходят 2 протона убихинола и перекачиваются комплексом.
-Комплекс IV (Цитохром c оксидаза) катализирует перенос 4 электронов с 4 молекул цитохрома на O2 и перекачивает при этом 4 протона в межмембранное пространство. Комплекс состоит из цитохромов a и a3, которые, помимо гема, содержат ионы меди.
Кислород, поступающий в митохондрии из крови, связывается с атомом железа в геме цитохрома a3 в форме молекулы O2. Каждый
из атомов кислорода присоединяет по два электрона и два протона и превращается в молекулу воды.
Суммарное уравнение реакции гидроксилирования вещества RH ферментами микросомального окисления:
RH + О2 + NADPH + Н+ → ROH + Н2О + NADP+.
Субстратами Р450 могут быть многие гидрофобные вещества как экзогенного, так и эндогенного происхождения.
в результате первой фазы обезвреживания с участием цитохрома Р450 происходит модификация веществ с образованием
функциональных групп, повышающих растворимость гидрофобного соединения. В результате модификации возможна потеря молекулой
её биологической активности или даже формирование более активного соединения, чем вещество, из которого оно образовалось.
ЦИТОХРОМ Р-450
Цитохром P450 — общее название ферментов семейства P450. Входят в класс гемопротеинов, относятся к цитохромам типа b.
Цитохром P450, связанный с монооксидом углерода, имеет максимум поглощения света при длине волны 450 нм, что определило его название.
Цитохромы P450 обнаружены во всех без исключения царствах живых существ — у животных, растений, грибов, бактерий, архей. У эукариотических организмов P450 являются мембранными белками.
Система цитохрома P450 участвует в окислении многочисленных соединений, как эндогенных, так и экзогенных. Ферменты этой группы играют важную роль в обмене стероидов, желчных кислот, ненасыщеных жирных кислот, а также в нейтрализации ксенобиотиков (лекарств, ядов, наркотиков)
Цитохром Р450-зависимые монооксигеназы катализируют расщепление различных веществ с участием донора электрона НАДФН и молекулярного кислорода. В этой реакции один атом кислорода присоединяется к субстрату, а второй восстанавливается до воды.
Ферменты семейства цитохрома P450, в отличие от остальных гемопротеинов, как правило, обладающих одним типом активности и строго определённой функцией, достаточно разнообразны по функциям, типам ферментативной активности, зачастую обладают малой субстратной специфичностью. P450 могут проявлять как монооксигеназную, так и оксигеназную активность, поэтому иногда относятся к оксидазам со смешанной функцией.
Оксигеназные реакции, катализируемые цитохромом Р450, весьма разнообразны. Цитохромы P450 катализируют омега-окисление насыщенных жирных кислот, перекисное окисление ненасыщенных жирных кислот, гидроксилирование стероидных гормонов, желчных кислот и холестерина, биосинтез простагландинов и другие реакции.
ГЛЮКОЗОЛАКТОННЫЙ ЦИКЛ
Глюкозо-лактатный цикл – это циклический процесс, объединяющий реакции глюконеогенеза и реакции анаэробного гликолиза.
Лактат, образованный в анаэробном гликолизе, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват. Лактат как источник пирувата важен не столько при голодании, сколько при нормальной жизнедеятельности организма. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата.
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NADH/NAD+ ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном направлении, т.е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют "глюкозо-лактатным циклом", или "циклом Кори". Цикл Кори выполняет 2 важнейшие функции: 1 - обеспечивает утилизацию лактата; 2 - предотвращает накопление лактата и, как следствие этого, опасное снижение рН (лактоацидоз). Часть пирувата, образованного из лактата, окисляется печенью до СО2 и Н2О. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза.
ГЛЮКОЗОАЛАНИНОВЫЙ ЦИКЛ
Целью глюкозо-аланинового цикла также является уборка пирувата, но, кроме этого решается еще одна немаловажная задача – уборка лишнего азота из мышцы.
При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты трансаминируются с α-кетоглутаратом.
Полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой азота и пирувата из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа передается на синтез мочевины, пируват используется для синтеза глюкозы.
Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах → пируват в мышцах → аланин в мышцах → аланин в печени → глюкоза в печени → глюкоза в мышцах. Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда мышечные белки распадаются и многие аминокислоты используются в качестве источника энергии, а их азот необходимо доставить в печень.
Билет№9
ВИТАМИН В2
Рибофлавин (лактофлавин, витамин B2) — один из наиболее важных водорастворимых витаминов, кофермент многих биохимических процессов.
БИОЛОГИЧЕСКАЯ РОЛЬ
Рибофлавин является биологически активным веществом, играющим важную роль в поддержании здоровья человека. Биологическая роль рибофлавина определяется вхождением его производных флавинмононуклеотида (FMN) и флавинадениндинуклеотида (FAD) в состав большого числа важнейших оксилительно-восстановительных ферментов в качестве коферментов.
Флавиновые ферменты принимают участие в окислении жирных, янтарной и других кислот; инактивируют и окисляют высокотоксичные альдегиды, расщепляют в организме чужеродные D-изомеры аминокислот, образующиеся в результате жизнедеятельности бактерий; участвуют в синтезе коферментных форм витамина B6 и фолацина; поддерживают в восстановленном состоянии глутатион и гемоглобин.
ИСТОЧНИК
Печень и почки,дрожжи,яйца,миндаль,шампиньоны,белые грибы,лисички,творог,брокколи,белокочанная капуста,гречневая крупа,молоко,мясо,очищенный рис,макаронные изделия,белый хлеб, большинство фруктов и овощей
КОФЕРМЕНТНАЯ Ф-Я
В ферментах коферменты функционируют как промежуточные переносчики электронов и протонов, отщепляемых от окисляемого субстрата.
2. ЦТК - центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии — АТФ.
РЕАКЦИИ

БИОХИМИЧЕСКОЕ ЗНАЧЕНИЕ
1.Интегративное – цикл Кребса объединяет пути катаболизма углеводов, белков и жиров, т.к. в нем утилизируется молекулы ацетилКоА, образующиеся при расщеплении этих веществ.
2.Энергетическое. При расщеплении 1 молекулы ацетилКоА до конечных продуктов (СО2 и Н2О) генерируется 12 молекул АТФ.
3.Амфиболическое (двойственное). В ЦТК происходит не только катаболические процессы – окисление ацетилКоА. Субстраты ЦТК используются и для реакций синтеза (анабо лические процессы). Так, из оксалоацетата синтезируется аспарагиновая кислота; из альфа кетоглутаровой кислоты – глутаминовая; из оксалоацетата фосфоэнолпируват.
АНАПЛЕРОТИЧЕСКИЕ РЕАКЦИИ
Это реакции клеточного метаболизма, повышающие концентрацию субстратов ЦТК, образуя их в других метаболических путях.
Например:
1.Образование альфа-кетоглутарата и оксалоацетата в реакциях трансаминирования аминокислот;
2.Образование альфа–кетоглутарата в глутаматдегидрогеназной реакции;
3.Образование оксалоацетата из пирувата в пируваткарбоксилазной реакции.
АМФИБОЛИЧЕСКИЕ РЕАКЦИИ
Обратимые участки метаболических путей, состоящие из равновесных реакций и используемые организмами как для синтеза, так и для расщепления сложных соединений, называются амфиболическими.
ГИПЕРГЛИКЕМИЯ
При недостаточности содержания инсулина возникает заболевание, которое носит название «сахарный диабет»: повышается концентрация глюкозы в крови (гипергликемия) и уменьшается содержание гликогена в печени. Мышечная ткань при этом утрачивает способность утилизировать глюкозу крови. В печени при общем снижении интенсивности биосинтетических процессов: биосинтеза белков, синтеза жирных кислот из продуктов распада глюкозы – наблюдается усиленный синтез ферментов глюконеогенеза. Развитие гипергликемии при диабете можно рассматривать также как результат возбуждения метаболических центров в ЦНС импульсами с хеморецепторов клеток, испытывающих энергетический голод в связи с недостаточным поступлением глюкозы в клетки ряда тканей.
Гипергликемия может возникнуть не только при заболевании поджелудочной железы, но и в результате расстройства функции других эндокринных желез, участвующих в регуляции углеводного обмена. Так, гипергликемия может наблюдаться при гипофизарных заболеваниях, опухолях коркового вещества надпочечников, гиперфункции щитовидной железы.
РЕГУЛЯЦИЯ СОДЕРЖАНИЯ ГЛЮКОЗЫ В КРОВИ
Поддержание содержания глюкозы в крови на определенном уровне является примером одного из самых совершенных механизмов гомеостаза, в функционировании которого участвуют печень, внепеченочные ткани и некоторые гормоны. Глюкоза легко проникает в клетки печени и относительно медленно в клетки внепеченочных тканей. Следовательно, прохождение через клеточную мембрану является лимитирующей скорость стадией при потреблении глюкозы внепеченочными тканями. Поступившая в клетки глюкоза быстро фосфорилируется при действии гексокиназы. С другой стороны, вполне возможно, что более значительное влияние на потребление глюкозы печенью или на выход глюкозы из этого органа оказывают активность некоторых других ферментов и концентрации ключевых промежуточных продуктов. Тем не менее концентрация глюкозы в крови является важным фактором, регулирующим скорость потребления глюкозы как печенью, так и внепеченочными тканями.
АЛИМЕНТАРНАЯ ГИПЕРГЛИКЕМИЯ
Развивается при приеме больших количеств сахара. Этот вид гипергликемии используют для оценки состояния углеводного обмена (так называемая сахарная нагрузка). У здорового человека после одномоментного приема 100—150 г сахара содержание глюкозы в крови нарастает, достигая максимума — 1,5—1,7 г/л (150—170 мг%) через 30—45 мин. Затем уровень сахара крови начинает падать и через 2 ч снижается до нормы (0,8—1,2 г/л), а через 3 ч оказывается даже несколько сниженным
ГЛЮКОЗУРИЯ
Глюкозурия — наличие глюкозы в моче. В норме моча не содержит глюкозы, поскольку почки способны реабсорбировать (возвращать в кровоток) весь объём глюкозы, прошедший через почечный клубочек в просвет канальцев нефрона. В подавляющем большинстве случаев глюкозурия является сипмтомом декомпенсированного сахарного диабета как результат патологического увеличения концентрации глюкозы в крови. Редким исключением является нарушение реабсорбции в самой почке, — т. н. ренальная (почечная) глюкозурия. Глюкозурия ведёт к избыточной потере воды с мочой — дегидратации организма, развивающейся из-за усиления осмотического компонента диуреза.
Билет 10
1. Витамин PP существует в двух формах - никотиновой кислоты и никотиномида. Активное воздействие витамина PP на обменные процессы обусловлено его вхождением в состав ниацинамидадениндинуклеотида (НАД) и ниацинамидадениндинуклеотида фосфата (НАДФ), являющихся кофакторами ряда ферментов. Никотиновая кислота в организме входит в состав NAD и NADP, выполняющих функции коферментов различных дегидрогеназ (см. раздел 2). Синтез NAD в организме протекает в 2 этапа:
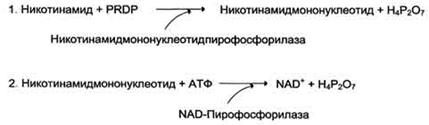
NADP образуется из NAD путём фосфорилирования под действием цитоплазматической NAD-киназы.
NAD+ + АТФ → NADP+ + АДФ
В частности, ниацинамид входит в состав кодегидраз, являющихся переносчиками водорода к флавопротеиновым ферментам, и тем самым регулирует окислительно-восстановительные процессы в организме. Он участвует более чем в полусотне реакций, в ходе которых сахар и жир превращаются в энергию. Он также необходим для обмена аминокислот и участвует в превращении жиров в вещества, именуемые эйкозаноидами, - гормоноподобные агенты, управляющие метаболическими путями нашего организма. Ниацин - витамин, не знающий равных в контроле холестерина. Ниацин борется с такими факторами риска сердечно-сосудистых заболеваний
Ниацин это белый кристаллический порошок без запаха, слабокислого вкуса. Трудно растворим в холодной воде (1:70), лучше в горячей (1:15), мало растворим в этаноле, очень мало — в эфире.Суточная потребность взрослого человека 15—20 мг.
Источники. Витамин РР широко распространён в растительных продуктах, высоко его содержание в рисовых и пшеничных отрубях, дрожжах, много витамина в печени и почках крупного рогатого скота и свиней. Витамин РР может образовываться из триптофана (из 60 молекул триптофана может образоваться 1 молекула никотинамида), что снижает потребность в витамине РР при увеличении количества триптофана в пище.
Недостаточность витамина РР приводит к заболеванию "пеллагра", для которого характерны 3 основных признака: дерматит, диарея, деменция ("три Д"), Пеллагра проявляется в виде симметричного дерматита на участках кожи, доступных действию солнечных лучей, расстройств ЖКТ (диарея) и воспалительных поражений слизистых оболочек рта и языка. В далеко зашедших случаях пеллагры наблюдают расстройства ЦНС (деменция): потеря памяти, галлюцинации и бред.
2. Гликоген служит в животном организме резервом углеводов, из которого по мере метаболической потребности могут высвобождаться глюкозофосфат или глюкоза. Хранение в организме собственно глюкозы неприемлемо из-за ее высокой растворимости: высокие концентрации глюкозы создают в клетке высоко гипертоническую среду, что приводит к притоку воды. Напротив, нерастворимый гликоген осмотически почти неактивен.
Гликоген животных, как и амилопектин растений, представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены α(1→4)-гликозидной связью. Связи в точках ветвления находятся в положении α(1→6) примерно каждого 10-го остатка. Таким образом, возникает древовидная структура с молекулярной массой >1ּ107 Да (до 50 000 остатков), в которой имеется только одна свободная аномерная ОН-группа, т. е. только один восстанавливающий конец. 
Синтез и распад гликогена. 1 - гексокиназа или глюкокиназа (печень); 2 - УДФ-глюкопирофосфорилаза; 3 - гликогенсинтаза; 4 - амило-1,4 → 1,6-глюкозилтрансфераза (фермент ветвления); 5 - гликогенфосфорилаза; 6 - "деветвящий" фермент; 7 - глюкозо-6-фосфатаза (печень); 8 - транспортные системы ГЛЮТ.
Гликоген печени никогда не расщепляется полностью. Как правило, укорачиваются или удлиняются (при высоком содержании глюкозы) только невосстанавливающие концы древовидной структуры. Удлинение цепи катализируется гликоген-синтазой [2]. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом [УТФ (UTP)] образуется активированный предшественник — УДФ-глюкоза (UDP-глюкоза) ([1]). После этого остаток глюкозы легко переносится с этого промежуточного соединения на гликоген. Когда растущая цепь достигает определенной длины (>11 остатков), специальный фермент ветвления гликогена (1,4→1,6-трансгликозидаза) [3] катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи] Дальнейшее удлинение этого фрагмента осуществляется гликоген-синтазой, образующей α(1→4)-связи.
Разветвленная структура гликогена облегчает быстрое освобождение углеводных остатков. Наиболее важным ферментом деградации гликогена является гликоген-фосфорилаза [4], отщепляющая от невосстанавливающего конца цепи остатки глюкозы в виде глюкозо-1-фосфата. Чем больше таких концов, тем больше молекул фосфорилазы могут действовать одновременно. Образование глюкозо-1-фосфата вместо глюкозы имеет то преимущество, что для включения освобожденных остатков глюкозы в гликолиз или ГМП не требуется АТФ.Благодаря структуре гликоген-фосфорилазы, процесс последовательного отщепления останавливается за 4 остатка глюкозы от точки разветвления. Точки ветвления удаляются двумя другими ферментами [5 и 6]. Вначале трисахарид боковой цепи переносится [5] к невосстанавливающему концу главной цепи. Затем 1,6-гликозидаза [6] отщепляет остающийся единичный остаток глюкозы в точке ветвления в виде свободной глюкозы, после чего неразветвленная цепь, может вновь расщепляться фосфорилазой.
В организме человека может содержаться до 450 г гликогена, треть из которого накапливается в печени, а остальное — главным образом в мышцах. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови в фазе пострезорбции. Поэтому содержание гликогена в печени варьирует в широких пределах. При длительном голодании оно падает почти до нуля, после чего начинается снабжение организма глюкозой с помощью глюконеогенеза. Гликоген мышц служит резервом энергии и не участвует в регуляции уровня глюкозы в крови. В мышцах отсутствует глюкозо-6-фосфатаза, поэтому гликоген мышц не может быть источником глюкозы в крови. По этой причине колебания содержания гликогена в мышцах меньше, чем в печени.
Фосфоролиз является основным путем распада гликогена, его катализирует фермент гликогенфосфорилаза, относящийся к классу трансфераз. Гликогенфосфорилаза отщепляет остатки глюкозы с нередуцирующего конца гликогена и переносит их на молекулу фосфорной кислоты с образованием глюкозо-1-фосфата:

Глюкозо-1-фосфат быстро изомеризуется, превращаясь в глюкозо-6-фосфат, который в печени гидролизуется фосфатазами до глюкозы и фосфорной кислоты:

Процесс фосфоролиза гликогена тонко регулируется. Регуляция активности гликогенфосфорилазы носит каскадный характер, в котором можно выделить несколько видов регуляции ферментативной активности:
1) гормональная (глюкагон в печени, адреналин в мышцах);2) аллостерическая;3) протеинкиназные реакции (в данном случае - фосфорилирование бокового радикала серина в гликогенфосфорилазе).
Активность мышечной фосфорилазы увеличивается при определенной концентрации АМФ и ацетилхолина, а также в присутствии катионов кальция и натрия.Снижение скорости фосфоролиза происходит при уменьшении запасов гликогена и фосфорной кислоты, а также при увеличении концентрации глюкозо-6-фосфата. Механизмы, снижающие скорость фосфоролиза гликогена, предохраняют организм от больших трат углеводных запасов (гликогена), которые могли бы привести к недостатку глюкозы, необходимой для работы головного мозга и сердечной мышцы.
3. Клеточная мембрана - это неотъемлемый компонент любой клетки. Ее роль в первую очередь состоит в том, чтобы отграничить внутреннее пространство клетки от внешней среды, а у эукариот, кроме того, разделить внутреннюю часть клетки на функционально значимые отсеки: ядро и митохонодрии. В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды. Наиболее важная группа, фосфолипиды, включает фосфатидилхолин (лецитин), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит и сфингомиелин. Холестерин присутствует во внутриклеточных мембранах животных клеток (за исключением внутренней мембраны митохондрий). Гликолипиды входят в состав многих мембран (например, во внешний слой плазматических мембран). В состав гликолипидов входят углеводные функциональные группы которые ориентируются в водную фазу.
Липиды мембран представляют собой амфифильные молекулы с полярной гидрофильной головкой (голубого цвета) и неполярным липофильным хвостом (желтого цвета). В водной среде они агрегируют за счет гидрофобных взаимодействий и вандерваальсовых сил.
Даниелли в связи с необходимостью объяснить явное расхождение между поверхностным натяжением на границах раздела масло/вода и мембра-на/вода. Была высказана гипотеза, что мембрана состоит из двойного липидного слоя, и предположено, что белок располагается на ее поверхности – модель Даниели – Дэвсона, или модель «сэндвича»). Это была очень удачная модель, и в течение последующих 30 лет многочисленные экспериментальные данные, особенно полученные с помощью дифракции рентгеновских лучей и электронной микроскопии, полностью подтвердили ее адекватность. Основными компонентами биологической мембраны являются липид и белок, вопрос о взаимном расположении этих компонентов в мембране стал предметом многочисленных дискуссий, так как обнаружилось, что мембраны выполняют разнообразные функции.
ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ МЕМБРАН КЛЕТКИ
Данная модель основана на предшествующих моделях структурно-функциональной организации мембран клетки. Живаямембрана представляет собой двумерный раствор глобулярных интегральных белков, диспергированных в жидкомфосфолипидном матриксе. Экспериментальные подтверждения данного предположения были получены при искусственно вызванном слиянии двух разных родительских клеток. При образовании плазматической мембраны гибридной клетки происходит быстрое стохастическое перемещение с систематическим упорядочением видоспецифичных белков и фосфолипидов. Такие перемещения в плоскости мембраны были названы латеральной подвижностью (диффузией) компонентов мембран.
Билет 11
1. Витамин В6(пиридоксин, пиридоксаль,
пиридоксамин)
В основе структуры витамина В6 лежит пиридиновое кольцо. Известны 3 формы витамина В6, отличающиеся строением замещающей группы у атома углерода в п-положении к атому азота. Все они характеризуются одинаковой биологической активностью.
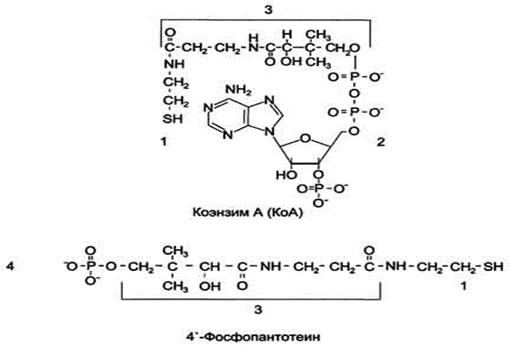
Рис. 3-1. Строение КоА и 4'-фосфопантотеина. 1 - тиоэтаноламин; 2 - аденозил-3'-фосфо-5'-дифосфат; 3 - пантотеновая кислота; 4 - 4'-фосфопантотеин (фосфорилированная пантотеновая кислота, соединённая с тиоэтаноламином).
Все 3 формы витамина - бесцветные кристаллы, хорошо растворимые в воде.
Источники витамина В6 для человека - такие продукты питания, как яйца, печень, молоко, зеленый перец, морковь, пшеница, дрожжи. Некоторое количество витамина синтезируется кишечной флорой.
Суточная потребность составляет 2-3 мг.
Биологические функции. Все формы витамина В6 используются в организме для синтеза кофер-ментов: пиридоксальфосфата и пиридоксаминфосфата. Коферменты образуются путём фос-форилирования по гидроксиметильной группе в пятом положении пиримидинового кольца при участии фермента пиридоксалькиназы и АТФ как источника фосфата.
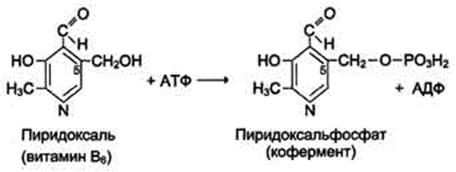
Пиридоксалевые ферменты играют ключевую роль в обмене аминокислот: катализируют реакции трансаминирования и декарбоксилирования аминокислот, участвуют в специфических реакциях метаболизма отдельных аминокислот: серина, треонина, триптофана, серосодержащих аминокислот, а также в синтезе тема (см. разделы 9, 12).
Клинические проявления недостаточности витамина. Авитаминоз В6 у детей проявляется повышенной возбудимостью ЦНС, периодическими судорогами, что связано, возможно, с недостаточным образованием тормозного медиатора ГАМК (см. раздел 9), специфическими дерматитами. У взрослых признаки гиповитаминоза В6 наблюдают при длительном лечении туберкулёза изониазидом (антагонист витамина В6). При этом возникают поражения нервной системы (полиневриты), дерматиты.
2. Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи). Следует отметить, что синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т.е. требующим затрат энергии. Глюкоза, поступающая в клетку, фосфорилируется при участии АТФ (реакция 1). Затем глюкозо-6-фосфат в ходе обратимой реакции превращается в глюкозо-1 -фосфат под действием фермента фосфоглюкомутазы. Глюкозо-1-фосфат по термодинамическому состоянию мог бы служить субстратом для синтеза гликогена. Но в силу обратимости реакции глюкозо-6-фосфат ↔ глюкозо-1-фосфат синтез гликогена из глюкозо-1-фосфата и его распад оказались бы также обратимыми и поэтому неконтролируемыми. Чтобы синтез гликогена был термодинамически необратимым, необходима дополнительная стадия образования уридинди-фосфатглюкозы из УТФ и глюкозо-1-фосфата. Фермент, катализирующий эту реакцию, назван по обратной реакции: УДФ-глюкопирофосфорилаза. Однако в клетке обратная реакция не протекает, потому что образовавшийся в ходе прямой реакции пирофосфат очень быстро расщепляется пирофосфатазой на 2 молекулы фосфата.
Реакция образования УДФ-глюкозы обусловливает необратимость всей серии реакций, протекающих при синтезе гликогена. Этим же объясняется невозможность протекания распада

Образование УДФ-глюкозы.

Образованная УДФ-глюкоза далее используется как донор остатка глюкозы при синтезе гликогена. Эту реакцию катализирует фермент гликогенсинтаза (глюкозилтрансфераза). Поскольку в данной реакции не используется АТФ, фермент называют син-тазой, а не синтетазой. Нуклеотидная часть УДФ-глюкозы играет существенную роль в действии гликоген синтазы, выполняя функцию "рукоятки", при помощи которой фермент располагает глюкозу в полисахаридной цепи в нужном положении. Кроме того, нуклеотидная часть УДФ-глюкозы, по-видимому, необходима для узнавания субстрата при катализе.
Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида, называемой "затравка" или "праймер". К "затравке" последовательно присоединяются молекулы глюкозы. Строением молекулы "затравки" как бы предопределяется тип связи, который возникает в реакции трансгли-козилирования. Таким образом, синтезируется полисахарид, аналогичный по строению с "затравочным". В состав "затравки" может входить белок гликогенин, в котором к ОН-группе одного из тирозиновых остатков присоединена олигосахаридная цепочка (примерно 8 остатков глюкозы). Глюкозные остатки переносятся гликогенсинтазой на нередуцирующий конец олигосахарида и связываются α-1,4-гликозидными связями. По окончании синтеза гликогенин остаётся включённым в гранулу гликогена.
Разветвлённая структура гликогена образуется при участии амило-1,4 →1,6-глюкозилтрансферазы, называемой ферментом "ветвления" (от англ, branching enzyme). Как только гликогенсинтаза удлиняет линейный участок примерно до 11 глюкозных остатков, фермент ветвления переносит её концевой блок, содержащий 6-7 остатков, на внутренний остаток глюкозы этой или другой цепи. В точке ветвления концевой остаток глюкозы олигосахарида соединяется с гидроксильной группой в С6 положении с образованием α-1,6-гликозидной связи. Новая точка ветвления может быть образована на расстоянии не менее 4 остатков от любой уже существующей. Таким образом, по мере синтеза гликогена многократно возрастает число ветвлений. Концы цепей служат точками роста молекулы при её синтезе и началом при её распаде.
Активность гликогенсинтазы также изменяется в результате фосфорилирования и дефосфорилирования (см. выше рис. 7-27). Однако есть существенные различия в регуляции гликогенфосфорилазы и гликогенсинтазы:
- фосфорилирование гликогенсинтазы катализирует ПК А и вызывает её инактивацию;
- дефосфорилирование гликогенсинтазы под действием фосфопротеинфосфатазы, наоборот, её активирует.

3. Аминокислот — эт органически соединения, физико-химическо по
ведени и разнообразны реакци которы объясняютс одновременны
присутствие в молекул основно аминогрупп N H2— и кисло карбо
ксильно групп —СООН.
3 Существуют три источника аминокислот в клетке – поступление из крови, распад собственных внутриклеточных белков и синтез заменимых аминокислот.
Путь дальнейшего превращения аминокислот зависит от вида и функции клетки, условий ее существования и гормональных влияний. Спектр веществ, получаемых клеткой из аминокислот, чрезвычайно широк.

Возможные пути превращений аминокислот
Значение белков для организма:
1. Как известно, белки представляют собой высокомолекулярные органические вещества, являющиеся основным структурным элементом всех клеток и тканей, пластическим субстратом для роста и развития организма, процессов регенерации. Недостаток белков ведет к алиментарной дистрофии, выражающейся в похудании, так как организм человека не может синтезировать белки из неорганических веществ и начинает расщеплять собственные белки, в частности белки скелетной мускулатуры. Дефицит белка приводит к замедлению роста и развития в детском и юношеском возрасте.
2. Белки являются ферментами и гормонами, катализируя обменные процессы и выполняя регуляторную функцию. Таким образом, при недостатке белков нарушается нормальное течение обменных процессов.
3. Иммуноглобулины (антитела) являются белками и выполняют защитную функцию. Значительный дефицит белка может привести к имму-нодепрессии, снижению реактивности и резистентности организма.
4. Белок имеет большое значение в деятельности центральной нервной системы. Недостаток белка в пище приводит к снижению внимания, работоспособности и тд.
5. Недостаток белка в пище приводит к понижению барьерной функции печени, изменениям эндокринной системы.

