




Окислительное фосфорилирование — один из важнейших компонентов клеточного дыхания, приводящего к получению энергии в виде АТФ. Субстратами окислительного фосфорилирования служат продукты расщепления органических соединений — белки, жиры и углеводы. Однако чаще всего в качестве субстрата используются углеводы.
Процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — Протонная АТФ-синтаза, состоящая из 9 субъединиц 5 типов. На внутренней мембране митохондрий также находится переносчик Фосфата из цитоплазмы в митохондрию, но для его работы необходим протон водорода. Такие переносчики называются транслоказами.
Ингибиторы блокируют V комплекс:
- Олигомицин — блокируют протонные каналы АТФ-синтазы.
- Атрактилозид, циклофиллин — блокируют транслоказы.
Разобщители — липофильные вещества, которые способны принимать протоны и переносить их через внутреннюю мембрану митохондрий минуя V комплекс (его протонный канал). Разобщители:
Естественные — продукты перекисного окисления липидов, жирных кислот с длинной цепью; большие дозы тиреоидных гормонов.
Искусственные — динитрофенол, эфир, производные витамина К, анестетики.
3.ГЛЮКОНЕОГЕНЕЗ
— процесс образования в печени и отчасти в корковом веществе почек (около 10 %) молекул глюкозы из молекул других органических соединений — источников энергии, например свободных аминокислот, молочной кислоты, глицерина. Свободные жирные кислоты у млекопитающих для глюконеогенеза не используются.
Глюконеогенез: субстраты, ферменты, регуляция и биологическая роль процесса.
Глюконеогенез - процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Это объясняется тем, что клетки мозга не способны, в отличие от других тканей, обеспечивать потребности в энергии за счёт окисления жирных кислот.
Кроме мозга, в глюкозе нуждаются ткани и клетки, в которых аэробный путь распада невозможен или ограничен, например эритроциты (они лишены митохондрий), клетки сетчатки, мозгового слоя надпочечников и др.
Первичные субстраты глюконеогенеза - лактат, аминокислоты и глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
- Лактат - продукт анаэробного гликолиза. Он образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
- Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке.
- Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза и катализируется теми же ферментами. Однако 3 реакции гликолиза термодинамически необратимы. На этих стадиях реакции глюконеогенеза протекают другими путями.
Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях.
1. Образование фосфоенолпирувата
из пирувата - первая из необратимых
стадий глюконеогенеза
Образование фосфоенолпирувата из пирувата происходит в ходе двух реакций, первая из которых протекает в митохондриях. Пируват, образующийся из лактата или из некоторых аминокислот, транспортируется в матрикс митохондрий и там карбоксилируется с образованием оксалоацетата.
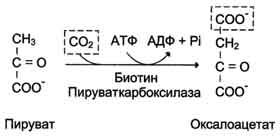
Пируват-карбоксилаза, катализирующая данную реакцию, - митохондриальный фермент, коферментом которого является биотин. Реакция протекает с использованием АТФ.
Дальнейшие превращения оксалоацетата протекают в цитозоле. Следовательно, на этом этапе должна существовать система транспорта оксалоацетата через митохондриальную мембрану, которая для него непроницаема. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием маната
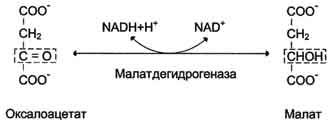 при участии NADH (обратная реакция цитратного цикла). Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспартатного челночного механизма.
при участии NADH (обратная реакция цитратного цикла). Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспартатного челночного механизма.
В цитозоле малат вновь превращается в оксалоацетат в ходе реакции окисления с участием кофермента NAD+. Обе реакции: восстановление оксалоацетата и окисление малага катализируют малатдегидрогеназа, но в первом случае это митохондриальный фермент, а во втором - цитозольный. Образованный в цитозоле из ма-лата оксалоацетат затем превращается в фосфоенолпируват в ходе реакции, катализируемой фосфоенолпируваткарбоксикиназой - ГТФ-зависимым ферментом.
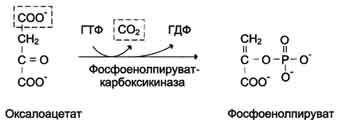
В пересчёте на синтез одной молекулы глюкозы из двух молекул пирувата расход составляет 2 моль АТФ и 2 моль ГТФ или 4 моль АТФ (для удобства рассуждений предлагается считать, что энергозатраты на синтез АТФ и ГТФ равны). После образования фосфоенолпирувата все остальные реакции также протекают в цитозоле вплоть до образования фруктозо-1,6-бисфосфата и катализируются гликолитическими ферментами.
2. Гидролиз фруктозо-1,6-бисфосфата
и глюкоза-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфата - также необратимые реакции глюконеогенеза. В ходе гликолиза эти реакции катализируют специфические киназы с использованием энергии АТФ. В глюконеогенезе они протекают без участия АТФ и АДФ и ускоряются не киназами, а фосфатазами - ферментами, принадлежащими к классу гидролаз. Ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза катализируют отщепление фосфатной группы от фруктозо-1,6-бисфосфата и глюкозо-6-фосфата. После чего свободная глюкоза выходит из клетки в кровь.
Итак, в печени существуют 4 фермента, которые принимают участие только в глюконеогенезе и катализируют обходные реакции необратимых стадий гликолиза. Это - пируват-карбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозе-1,6-бисфосфатаза и глюкозо-6-фосфатаза.
Суммарный результат глюконеогенеза из пирувата выражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NADH + Н+)+ 4 Н20 → Глюкоза + 4 АДФ + 2 ГДФ + 6 H3PO4 + 2 NAD+
СУБСТРАТЫ
РЕАКЦИИ
РЕГУЛЯЦИЯ
Билет №8
ВИТАМИНЫ
Витамины – это низкомолекулярные органические соединения, необходимые для нормальной жизнедеятельности, и поступают в организм в незначительном количестве с продуктами питания.
КЛАССИФИКАЦИЯ
В зависимости от растворимости в неполярных органических растворителях или в водной среде различают жирорастворимые и водорастворимые витамины.
1)Водорастворимые витамины (аскорбиновая кислота,В1,В2,В16..) берут участие в обмене веществ как коферменты или составные ферментов.
2)Жирорастворимые витамины (А,Д,К,Е) не входят в состав ферментов и непосредственно влияют на обмен веществ, создавая условия для оптимального действия ферментов на мембранных структурах.
БИОЛОГИЧЕСКАЯ РОЛЬ
Часто витамины входят в состав ферментов, то есть биологических катализаторов процессов живой клетки.
СУТОЧНАЯ ПОТРЕБНОСТЬ
Концентрация витаминов в тканях и суточная потребность в них невелики, но при недостаточном поступлении витаминов в организм наступают характерные и опасные патологические изменения.
Большинство витаминов не синтезируются в организме человека. Поэтому они должны регулярно и в достаточном количестве поступать в организм с пищей или в виде витаминно-минеральных комплексов и пищевых добавок. Исключения составляют витамин К, достаточное количество которого в норме синтезируется в толстом кишечнике человека за счёт деятельности бактерий, и витамин В3, синтезируемый бактериями кишечника из аминокислоты триптофана.
ВИТАМИН В1
Тиами́н (витамин B1) — водорастворимый витамин, соединение, отвечающее формуле C12H17N4OS. Бесцветное кристаллическое вещество, хорошо растворимое в воде, нерастворимое в спирте. Разрушается при нагревании. Тиамин, являясь водорастворимым соединением, не запасается в организме и не обладает токсическими свойствами.
БИОЛОГИЧЕСКАЯ РОЛЬ
Тиамин играет важную роль в процессах метаболизма углеводов и жиров. Вещество необходимо для нормального протекания процессов роста и развития и помогает поддерживать надлежащую работу сердца, нервной и пищеварительной систем.
Гиповитаминоз В1. Нарушается окисление пировиноградной кислоты, так как витамин B1 входит в состав кофермента, участвующего в этом процессе. Пировиноградная кислота накапливается в избытке и частично переходит в молочную кислоту, содержание которой также возрастает. При нарушении окисления пировиноградной кислоты снижается синтез ацетилхолина и нарушается передача нервных импульсов. Уменьшается образование из пировиноградной кислоты ацетилкоэнзима А.
Пировиноградная кислота является фармакологическим ядом для нервных окончаний. При увеличении ее концентрации в 2—3 раза возникают нарушения чувствительности, невриты, параличи и др.
При гиповитаминозе B1 нарушается также и пентозофосфатный путь обмена углеводов, в частности образование рибозы.
ИСТОЧНИКИ
Основные количества тиамина человек получает с растительной пищей. Богаты тиамином такие растительные продукты, как пшеничный хлеб из муки грубого помола, соя, фасоль, горох, шпинат. Меньше содержание тиамина в картофеле, моркови, капусте. Из животных продуктов содержанием тиамина выделяются печень, почки, мозг, свинина, говядина, также он содержится в дрожжах. В молоке его содержится около 0,5 мг/кг.[5] Витамин B1 синтезируется некоторыми видами бактерий, составляющих микрофлору толстого кишечника.
КОФЕРМЕНТНАЯ Ф-Я
Витамин В1 выступает как кофермент ферментных систем организма, которые контролируют цепи углеводного, липидного и белкового обмена.
Витамин В1 принадлежит к числу витаминов, механизм действия которых достаточно ясен.
Было показано, что пирофосфорный эфир тиамина представляет собой кофермент карбоксилазы, а также дегидрогеназ, катализирующих окислительное декарбоксилирование кетокислот (пировиноградной и альфа-кетоглютаровой).

