




Субстратом для синтеза нуклеиновых кислот в клетках и тканях могут служить продукты гидролиза ДНК и РНК, а также продукты обмена белков и углеводов.
Биосинтез пуриновых нуклеотидов происходит из углекислого газа, глицина, муравьиной кислоты, аспарагиновой кислоты и глутамина. В результате поэтапного синтеза из этих компонентов (всего 11 реакций) образуется инозиновая кислота, из которой затем АМФ и ГМФ. Из указанных монофосфатов затем синтезируются соответствующие ди- и трифосфаты.
Синтез пиримидиновых нуклеотидов происходит также поэтапно, вначале образуется карбомоилфосфат из NH3, CO2, АТФ. Затем с участием аспарагиновой кислоты образуется дигидрооротовая кислота, затем оротовая кислота.
Оротовая кислота далее взаимодействует с 5'-фосфорибозил-1-пирофосфатом, при этом синтезируется оротидин-5 -фосфат, который затем декарбоксилируется с образованием уридинмонофосфата. Из уридиловой кислоты при аминировании углеродов за счет глутамина образуется цитидинмонофосфат, а при метилировании – тимидиловая кислота:

Эти монофосфаты затем фосфорилируются с участием АТФ до образования трифосфатов.
Синтез ДНК (или репликация ДНК). В клетках животных синтез (репликация) ДНК происходит только в определенный период жизни клетки – в синтетической S-фазе. Эта фаза отделена от митоза (M) предсинтетическим (G1) и постсинтетическим (G2) периодами (рис. 12.1).

Рис. 12.1. Цикл деления клеток млекопитающих.
Фаза синтеза ДНК (S-фаза) отделена от митоза (M) периодами G1 и G2. Стрелкой указано направление синтеза.
Репликация ДНК происходит в строго определенное время в клетках, готовящихся к делению. В S-фазе происходит полная и строго однократная репликация ядерной ДНК, по времени это занимает примерно 9 часов; за этот срок происходит удвоение генетического материала диплоидной делящейся клетки. Репликация начинается сразу на многих участках молекулы ДНК, называемых точками начала репликаци, обозначаемых ori (origin – начало). Биосинтез ДНК происходит с участием всех четырех дезоксирибонуклеозидтрифосфатов – АТФ, ГТФ, ЦТФ, ТТФ; ферментов синтеза ДНК: ДНК-полимераз, хеликазы – фермента, обеспечивающего раскручивание (расплетение) двойной спирали ДНК в репликативной вилке, топоизомеразы фермента, способствующего расщеплению спирали ДНК в области репликативной вилки, ферментов, «редактирующих» ДНК, т.е. осуществляющих вырезание и удаление ошибочно включенных нуклеотидов или репарирующих повреждения ДНК.
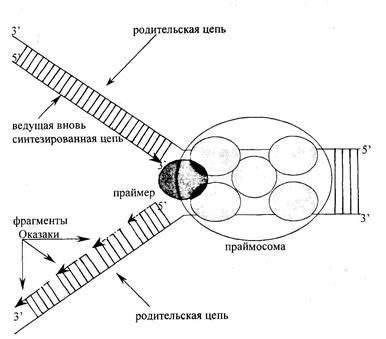
Все этапы синтеза ДНК эукариот осуществляются с помощью репликативного мультиферментного комплекса (праймосомы), состоящего из ферментов и белковых факторов, осуществляющих узнавание точки ori, расплетание родительского дуплекса, удержание материнских цепей ДНК на достаточном расстоянии друг от друга, инициацию новых дочерних цепей, закручивание цепей в спираль и, наконец, терминацию репликации ДНК. В составе комплекса присутствуют новосинтезированная ДНК, не менее 30 белков с Мм от 15 до 300 кДа, ДНК-полимеразы 5, а, праймаза, 3'-5'экзонуклеаза, ДНК-лигаза I, РНКаза H, ДНК-топоизомераза I, ДНК-хеликаза, белок PCNA (ядерный антиген пролиферирующих клеток) и другие факторы.
В праймасоме репликативный белок A (RPA) связывается с однонитевой ДНК и существует в виде комплекса из трех белков с Мм 70, 34 и 11 кДа. Репликативный фактор С (RFC) состоит из пяти белковых елиниц и играет важную роль при связывании репликативных полимераз во время образования репликативной вилки. Белок PCNA, так называемый ДНК-полимеразный зажим, функционирует как кофактор ДНК-полимеразы, образуя закрытое кольцо, окружающее ДНК с каналом в центре. Репликативный фактор С совместно с PCNA продвигается вдоль ДНК по репликативной вилке. С участием праймасомы, ферментов образуются новые полинуклеотидные цепи. Синтез новых цепей всегда идет в направлении от 5'-конца к 3'-концу. Поэтому на одной из ветвей репликативной вилки новая цепь наращивается непрерывно по мере раскручивания ДНК-матрицы, а на другой ветви образуются короткие фрагменты новой цепи ДНК, называемые фрагменты Оказаки; затем концы этих фрагментов соединяются между собой в результате действия фермента ДНК-лигазы (рис. 12.2.).
5'
Рис. 12.2. Схематическое изображение синтеза ДНК в репликативной вилке.
На одной ветви репликативной вилки нуклеотидная цепь синтезируется непрерывно, на другой – прерывисто, с образованием фрагментов Оказаки. Фрагменты Оказаки соединяются между собой в результате действия ДНК-лигазы. По мере роста новых цепей репликативная вилка перемещается по ДНК и в результате происходит репликация (удвоение) молекулы ДНК. Последовательности нуклеотидов новой цепи ДНК строго комплементарны последовательностям родительской цепи, благодаря этому соблюдается передача наследственной информации из поколения в поколение (см. рис.3.5.). Точность репликации очень высока – возможна одна ошибка на 10 трансферазных реакций.
Синтез ДНК на матрице РНК происходит под действием фермента ревертазы или обратной транскриптазы (РНК-зависимой ДНК-полимеразы), содержащейся в составе онковирусов. Доказано, что многие РНК-содержащие онкогенные вирусы (онковирусы) содержат этот фермент в составе покровных белков. При этом механизм синтеза ДНК на матрице РНК включает 3 стадии: на первом этапе фермент ревертаза синтезирует на матрице вирусной РНК комплементарную цепь ДНК, что приводит к формированию гибридной молекулы. Затем исходная молекула РНК отделяется от ДНК и только после этого на матрице ДНК синтезируется комплементарная цепь ДНК.
Биосинтез РНК происходит в клетках на матрице ДНК, т.е. синтезируемая РНК комплементарна к одной из цепей этой ДНК. Если при репликации ДНК происходит удвоение всей молекулы ДНК, то при синтезе РНК-транскрипции копия с молекулы ДНК снимается лишь с отдельных генов, с отдельных участков одной из цепей ДНК. Синтез различных форм РНК осуществляется с участием соответствующих РНК-полимераз, которые катализируют синтез информационной (и-РНК), рибосомальной (р-РНК) и транспортной (т-РНК) рибонуклеиновых кислот. В природе существует фермент РНК-зависимая РНК-полимераза, который обеспечивает в клетке репликацию РНК у РНК-содержащих вирусов. При этом РНК вируса выполняет две функции – роль носителя генетической информации (вместо ДНК) и информационной РНК. Вирусная РНК является одноцепочечной. Эту цепь обозначают «плюс»-тип РНК, а синтезируемую в присутствии РНК-полимеразы комплементарную ей цепь РНК «минус»-цепь. Поступившая в клетку вирусная РНК выполняет роль информационной РНК, связывается с рибосомами клетки - хозяина и обеспечивает образование фермента репликазы и белков вирусной оболочки. После этого с помощью фермента репликазы синтезируется вирусная РНК на матрице исходной РНК. В свою очередь молекула «минус»-РНК является матрицей для синтеза цепей «плюс»-РНК.

