




Опыление — это процесс переноса пыльцы пыльников тычинок на рыльце пестика. У растений различают два типа опыления, в зависимой от того, пыльца какого цветка попадает на рыльце пестика:
Самоопыление, или автогамия — пыльца с тычинок попадает на рыльце пестика того же цветка.
Перекрестное, или аллогамия, — пыльца с тычинок одного цветка попадает на пестик другого цветка. Первыми перекрестное опыление открыли Доббе (1750) и Мюллер (1751), однако они не продолжили своих исследований. Наибольший вклад в его изучение внесли И. Г. Кёльрёйтер и К. К. Шпренгель. При перекрестном опылении выделяются следующие его виды:
а) гейтоногамия,или соседнее опыление, — пыльца с тычинок одного цветка попадает на рыльце пестика другого цветка в пределах одного соцветия или растения
б) ксеногамия пыльца с тычинок цветка, расположенного на одном растении, попадает на пестик цветка другого растения, но того же вида;
в) гибридизация на рыльце пестика цветка попадает пыльца с цветков другого сорта или вида.
В зависимости от того, какие агенты участвуют в переносе пыльцы при перекрестном опылении. (абиотические — ветер, вода и биотические -насекомые и другие животные), различаются следующие способы опыления: анемофилия, или ветроопыление; гидрофилия — опыление водой энтомофилия — насекомоопыление; орнитофилия —опыление птицами; мирмекофилия-опыление муравьями. В процессе сопряженной эволюции (коэволюции) растения и агенты-опылители (биотические) приспособились к наилучшему исполнению этого процесса, а растения, опыляемые абиотическими факторами, — к этим факторам. Самоопыление способствует стабилизации вида, а в селекции очень важно для выведения чистых линий. Часто совершается до распускания цветков: у злаков в то время, когда соцветие находится во влагалище листа; у гороха, фасоли — в бутоне. В это время пыльца других растений в цветок попасть не может (но это не значит, что не произойдет оплодотворение, если другая, пыльца попадет на рыльце). Однако есть растения, у которых перекрестное опыление исключено. Многие растения образуют клейстогамные цветки, мелкие, невзрачные, вовсе не раскрывающиеся, в которых возможно только самоопыление (арахис, летние цветки лесных фиалок, кислица, сердечник).
Самоопыление в растительном мире — явление малораспространенное, но среди культурных растений самоопылителей достаточно много: ячмень, пшеница, фасоль, овес, томат, хлопчатник.
Однако самоопыление нельзя считать признаком примитивности. И тем не менее в процессе эволюции у растений сформировались приспособления, исключающие самоопыление.
1. Самостерильность (автостерильность) — неспособность пыльцы прорастать на рыльце пестика собственного цветка (рожь, рис, кукуруза, капуста и др.), а у плодовых — даже пыльцы того же сорта
2. Дихогамия неодновременное созревание в цветке пыльников и рылец.
3. Гетеростилия (разностолбчатостъ) — наличие у растений одного и того же вида двух или трех форм цветков, различающихся по длине столбиков и размещению тычинок
4. Раздельнополость цветков.
5. Двудомность растений, при котором самоопыление исключено.
Перекрестное опыление. В биологическом отношении более предпочтительно, так как приводит к новым комбинациям признаков у дочернего организма, вследствие чего он имеет более широкий диапазон приспособительных признаков. Однако оно в значительной степени зависит от наличия агентов (при опылении животными) или от благоприятных условий среды. Всем известно, что обильно цветущие сады не плодоносят, если во время цветения идут продолжительные дожди. В силу большей ценности перекрестное опыление получило и более широкое распространение. Все особенности растений, препятствующие самоопылению, способствуют перекрестному: самостерильность, гетеростилия, дихогамия, раздельнополость и раздельнодомность.
Как уже отмечалось, в процессе эволюции произошла довольно узкая специализация отдельных агентов, участвующих в опылении, что и учтено при названии способов опыления.
Энтомофилия. Широко распространена в природе: около 80 % растений опыляются насекомыми. У этих растений сформировались приспособления для привлечения насекомых. Наиболее существенные из них следующие,
1. Большое количество пыльцы, которая служит пищей для насекомых. Пыльцевые зерна снабжены различными шипами, выростами, бугорками, иногда их поверхность клейкая, зерна крупные (магнолия, пион, кувшинки, калужница).
2. Нектар, синтезируемый нектарниками..
3. Энтомофильные растения имеют у цветков яркоокрашенный венчик или венчиковидный околоцветник.
4. Запах насекомые различают лучше, чем цвет, причем на значительных расстояниях, и он служит им ориентиром. Опыление насекомыми нередко выражается сложнейшей зависимостью между ними и растениями, и эта связь становится столь тесной, что растение оказывается в полной зависимости от своего опылителя. Менее распространены другие способы зоофилии:
а) мирмекофилия — опыление муравьями, которые используют растительные выделения в качестве пищи или устраивают гнезда в органах растения. В тропиках, например, муравьи поселяются в полых стволах Cecropia adenopus, в полых междоузлиях Clerodendrum myrmecophilum, полых колючках акаций, в полостях клубневидных стеблей некоторых эпифитов (Myrmecodia echinata). Часто муравья не только опыляют растения (переползая с цветка на цветок в поисках пищи), но и защищают его от насекомых (обычно листогрызущих);
б) кантарофилия опыление жуками, впервые отмеченное Дилсом в 1916 г. Это явление широко распространено в тропической зоне. У жуков, как правило, грызущий ротовой аппарат и некоторые его части хорошо адаптировались к пережевыванию пыльцы. Узкой специализации, за исключением навозников, нет. Жуками опыляются некоторые зонтичные, калина, виктория;
в) миофилия — опыление двукрылыми (жужжалы, слепни.). Только мухи опыляют плющ. Цветки таких растений неглубокие, обычно светлые, но не яркие. Растения часто имеют ловушки (кирказон);
г) хироптерофилия — опыление растений летучими мышами. Эти растения распускают цветки обычно ночью, выделяют нектар, неприятный запах; цветки крупные, имеют большое количество пыльцы. Соцветия хорошо выделяются изкроны;
д) орнитофилия - опыление птицами, характерно для тропиков. Орнитофильные растения цветут днем. Цветки яркие, контрастные, зигоморфные, запаха не имеют, выделяют много нектара, тычиночные нити твердые. Опыление производят колибри, нектарницы, гавайские цветочницы, медоеды, мелкие попугаи.
Анемофилия) - ветроопыление, которое присуще 10 % покрытосеменных и всем голосеменным. К этим растениям относятся злаки, осоковые, большинство пальм, древесных растений (березовые, ивовые, буковые, шелковица, платан и т. д.). Морфологические особенности анемофилънъгх растений: мелкие невзрачные цветки с простым чашечковидным венчиком; сухая пыльца, легкая и мелкая, иногда с воздушными мешками и образующаяся в огромных количествах (в одной сережке лещины 4 млн пылинок, в мужской метелке кукурузы — около 50 млн). У березы, орехов, лещины, дуба мужские сережки повислые и также легко раскачиваются ветром. Цветение у них происходит ранней весной до распускания листьев или одновременно с листораспусканием. Рыльца пестиков или клейкие, или волосистые, Анемофильные растения обычно имеют раздельнополые соцветия, пыльца переносится на расстояние до 60—70 км.
Гидрофилия — приспособленность цветков некоторых водных растений к опылению с помощью воды. Этот способ — наиболее древний по происхождению. Нельзя считать, что все растущие в воде растения гидрофильны. Прежде всего много мелких самоопыляющихся растений могут цвести будучи погруженными в воду. В клейстогамных цветках, как известно, опыление происходит внутри цветка. В то же время все рдесты, колосовидные соцветия которых приподнимаются над водой, ветроопыляемы, а водяная лобелия энтомофильна. Опыление водой осуществляется у растений, цветки которых целиком погружены в воду (роголистники, наяда и др.).
12.Белки - высокомолекулярные соединения, образующие при гидролизе аминокислоты. В организме животных белков содержится до 40-50% и более на сухую массу.
Функции белков: 1.Строительная, структурная. Белки образуют клеточные мембраны в комплексе с липидами. 2.Каталитическая. 3.Двигательная. 4.Транспортная. 5.Защитная. 6.Гормрнальная. 7.Запасающая. 8.Опорная. 9.Рецепторная. 10.Рецепторная. 11.Регуляторная.
Первичная структура – порядок чередования аминокислот в полипептидной цепи (или цепях) и местоположение дисульфидных связей. Полипептидная цепь содержит на одном конце свободную аминогруппу (N-конец), на другом - карбоксильную группу (С-конец). За начало цепи принимается ее N-конец, именно отсюда начинается отсчёт аминокислот.
Вторичная структура - упорядоченное пространственное расположение отдельных участков полипептидной цепи без учета типа и конформации боковых радикалов амино-кислот. Она образуется за счет замыкания водородных связей между пептидными групп-пами. Вторичная структура представлена в основном такими регулярными структурами как α-спираль, складчатые слои (β-структура). Часть полипептидной цепи не имеет упоря-доченного строения, такие участки называют бесструктурными областями Третичная структура - пространственное расположение упорядоченных и аморфных участков в полипептидной цепи в целом, которое достигается за счет взаимодействия бо-ковых радикалов и зависит от их типа и конформации Третичная структура имеет прямое отношение к форме молекул белка, которая может быть различной: от шарообразной до нитевидной. В поддержании третичной структуры, ее закреплении принимают участие различные типы связей: -водородные, -дисульфидные, -ионные, -гидрофобные. Четвертичная структура - способ взаимного расположения в пространстве отдельных полипептидных цепей в молекуле, характер связей между ними. Каждая полипеп-тидная цепь в составе молекулы белка с четвертичной структурой называется прото-мером или субъединицей. Белки, имеющие четвертичную структуру–олигомерные.
Аминокислоты, входящие в состав белков.
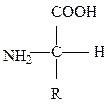
1. с неполярным Р.
глицин -Н
аланин -CH3
валин
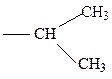
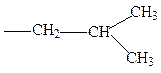
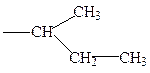
лейцин изолейцин
С полярным незаряженным Р
а) гидроксикислоты
серин
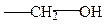
треонин
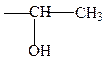
б) серосодержащие
цистеин метионин
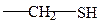
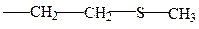
в) амиды дикарб. кислот
аспарагин
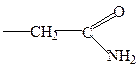
глутамин
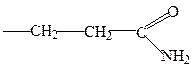
С ароматическим. Р
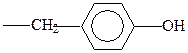
фенилаланин

тирозин
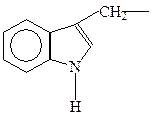 триптофан
триптофан
4. с “-“ заряженным Р
аспарагиновая
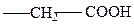 глутаминовая
глутаминовая 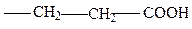
5. с “+” заряженным Р
лизин 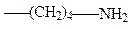
аргинин 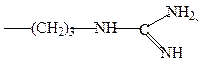
гистидин
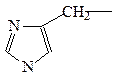 пролин (иминокислота)
пролин (иминокислота) 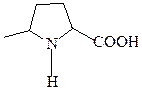
Денатурация белка – нарушение нативной пространственной структуры белковой молекулы, приводящее к утрате биологической активности и изменению физико-химических свойств. Денатурация не сопровождается разрывом пептидных связей. Происходит разрыв дисульфидных, гидрофобных, ионных, водородных связей, нарушается третичная и в значительной мере вторичная структура. Денатурацию вызывают химические соединения: орг. растворители, ионы тяжелых металлов, концентрированные кислоты и щелочи. Физические факторы: нагревание, высокое давление, ультрафиолетовое облучение.
Семейство Злаки
Мятликоцветные включают одно семейство Мятликовых — Роасеае. Иногда выделяют порядок Пленчатоцветных, куда относят также семейство Осоковых, конвергентно с ним сходное.
Семейство Мятликовых объединяет от 7500 до 10 000 видов (около 700 родов). Среди них есть виды-космополиты. Максимальное разнообразие жизненных форм приурочено к субтропическим и тропическим странам: деревья до 30—40м высоты, кустарники, но преимущественно многолетние и однолетние травы.

