




Еукаріотична клітина складається з: плазмолеми, цитоплазми і ядра. До цитоплазми належать: гіалоплазма, органели і включення.
1.2.1 Плазмолема, або зовнішня клітинна оболонка. Вона відмежовує клітину зовні і забезпечує її безпосередній зв'язок з позаклітинним се-редовищем. Вона утворена із трьох шарів: зовнішнього (глікокаліксу), середнього (клітинної мембрани), внутрішнього (підмембранного).
Середній шар складається з тонких (6-10 нм) пластів ліпопротеїдної природи, до складу яких входять 40% ліпідів, близько 60% білків, значна частина клітинних мембран містить 5-10% вуглеводів. Серед ліпідів зна-ходяться органічні речовини, які є гідрофобними — погано розчиняються у воді та характеризуються значною розчинністю в органічних розчин-никах та жирах — ліпофільні. Серед ліпідів, що входять до складу клі-тинних мембран, знаходяться фосфоліпіди, сфінгомієліни, холестерин. Особливістю ліпідних мембран є поділ молекул на дві функціонально різні частини: гідрофобні неполярні, що не несуть зарядів, «хвости», скла-даються із жирних кислот, та гідрофільні, заряджені полярні «голівки». Це визначає здатність ліпідів довільно утворювати двошарові (біліпідні) мембранні структури товщиною 5-7 нм. Клітинні мембрани можуть зна-чно різнитися між собою за ліпідним складом та білковими молекулами. Багато мембранних білків складаються з двох частин, з ділянок багатих полярними (що несуть заряд) амінокислотами, та ділянок, збагачених не-полярними амінокислотами. Такі білки в ліпідних шарах мембран розмі-щуються так, що їх неполярні ділянки ніби занурені в «жирну» частину мембрани, де знаходяться гідрофобні ділянки ліпідів (рис. 3).
Полярна (гідрофільна) частина цих білків взаємодіє з голівками ліпідів і обернена в бік водяної фази. Крім інтегральних білків, існують білки частково вмонтовані у мембрану — напівінтегральні та примемб-ранні, вмонтовані в біліпідний шар. За біологічним значенням білки мембран поділяються на білки-ферменти, переносники, рецепторні та структурні (рис. 4).
Зовнішній шар - глікокалікс (від лат. §1ікіз - солодкий + від гр. са-Іух — чашечка) має товщину 3-4 нм., містить вуглеводи олігоцукриди,
Розділ 1
Основи загальної цитології
які утворюють з білками і ліпідами клітинні мембрани, глікопротеї-ди та гліколіпіди. Ці структури забезпечують рецепторну функцію та здійснюють взаєморозпізнання та взаємодію клітин між собою.

Рис. 3. Схема молекулярної будови клітинної оболонки:
1-вуглеводні ланцюги; 2-гліколіпід; 3-глікопротеїд; 4-ліпідний хвіст; 5-по-лярна головка; 6-холестерин; 7-білок; 8-мікрофіламенти; 9-мікротрубочки.
Внутрішній або підмембранний шар, утворений мікрофіламентами та мікротрубочками. Ці структури забезпечуюють переміщення плаз-молеми, окремих ділянок клітини та клітини в цілому.
Плазмолема виконує функції розмежування цитоплазми із зо-внішнім середовищем, рецепції та транспорту різних речовин, серед яких важливе місце належить забезпеченню оптимального рівня аси-метрії концентрації іонів натрію і калію у клітині та за її межами, а та-кож забезпечує взаєморозпізнавання і взаємодію клітин з утворенням міжклітинних контактів, формування структури клітинної поверхні, рецепцію сигналізації з боку зовнішнього середовища (рис. 5).
Транспортна функція плазмолеми здійснюється шляхом дифузії па-сивного перенесення різних речовин — води, іонів, деяких низькомоле-кулярних сполук. Інші речовини проникають через мембрану шляхом активного перенесення проти градієнта концентрації за рахунок роз-щеплення АТФ. Так транспортуються цукри, амінокислоти. Ці процеси відбуваються за участю спеціальних білків — переносників. Транспорт із зовнішнього середовища в клітину називають ендоцитозом, за межі клі-
В.П. Новак, Ю.П. Бичков, М.Ю. Пшипенко Цитологія, гістологія, ембріологія
тини — екзоцитозом. При цьому внутрішньоклітинні продукти (білки, мукополісахариди, жирові краплі тощо) містяться у вакуолі або міхурці і відмежовані від гіалоплазми мембраною наближаються до плазмолеми. У місцях контакту плазмолема та мембрана вакуолі зливаються і вміст вакуолі виходить за межі клітини. Екзоцитоз поділяють на такі різнови-ди: секрецію — виділення клітиною продуктів її синтетичної діяльності; екскреція — виділення шкідливих продуктів метаболізму; клазматоз — видалення за межі клітин окремих її структурних компонентів.

Рис. 4. Гіпотетична схема плазматичної мембрани.
Ендоцитоз та екзоцитоз відбуваються за участю пов'язаних з плаз-молемою системи фібрилярних компонентів цитоплазми — мікро-трубочок та скорочувальних мікрофіламентів. Великі молекули та їх агрегати проникають у клітину шляхом фагоцитозу, який був вперше описаний І.І. Мечниковим.
Поглинання частинок рідини називають піноцитозом. Загальним для цих процесів є те, що поглинання на поверхні плазмолеми відбува-ється шляхом оточення речовини ділянкою плазматичної мембрани у вигляді вакуолі, яка переміщується у цитоплазму. Поглинуті частинки розщеплюються за допомогою ферментів і їхні складові засвоюються
Розділ 1
Основи загальної цитології
клітиною. Таким чином, поглинуті речовини у середині мембранних накуолей, утворених із елементів плазмолеми, підлягають внутрішньо-клітинному перетравленню. Процес, при якому поглинені частинки в оточенні мембрани проходять через цитоплазму і виводяться без змін за межі клітини називають цитопемпсисом.
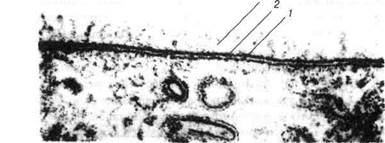
Рис. 5. Електронна мікрофотографія клітинної мембрани і глікокалікса х 200 000:
1-субмембранний комплекс; 2-плазматична мембрана; 3-глікокалікс
Рецепторні функції плазмолеми пов'язані з локалізацією на ній спеціальних структур, які беруть участь в специфічному пізнанні хімічних та фізичних факторів. Клітинна поверхня має велику кіль-кість рецепторів, що визначають можливість специфічних реакцій. Існують рецептори до біологічно-активних речовин — гормонів, ме-діаторів, антигенів тощо. Складні процеси рецепції є основою вза-єморозпізнання клітин, важливою і необхідною умовою існування багатоклітинних організмів.
Міжклітинні контакти
Міжклітинні контакти найбільше притаманні епітеліальним та ен-дотеліальним клітинам.
Розрізняють такі типи міжклітинних контактів:
Просте міжклітинне сполучення — плазмолеми сусідніх клітин на-ближаються одна до одної на відстань 15-20 нм. При цьому відбуваєть-ся взаємодія шарів глікокалікса. Розрізняють також щільне з 'єднання за типом замка, при цьому шари двох плазмолем наближені максималь-
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
но — ділянки плазмолем двох сусідніх клітин ніби зливаються. Ця ді-лянка непроникна для молекул та іонів. Такий тип з'єднання плазмолем називають десмосомою. Вона являє собою площу діаметром до 0,5 мкм, інколи має вигляд шарів; між мембранами знаходиться зона з високою електронною щільністю; зміцнення зв'язку між клітинами досягаєть-ся за допомогою фібрилярних структур цитоплазми та кортикально-го шару плазмолеми. Спостерігаються з'єднання типу напівдесмосом, якщо десмосома складається лише з однієї пластинки прикріплення.
Щільний замикаючий контакт характерний для апікальної поверх-ні клітин облямівкового епітелію, а також ендотелію, мезотелію. Такий

|
контакт характеризується максимальним зближенням плазматичних мембран сусідніх клітин, проміжок ущільнюється за рахунок анастомозуючих фібрил та іонів кальцію. При цьому зовнішні гідрофільні шари і глікокалікс суміжних плазмолем тісно зливаються в один суцільний шар завтовшки 2-3 нм. У серцевих м'язах зустрічаються щілинні контакти (не-ксуси), що забезпечують безпосередній обмін речовин між сусідніми клітинами. Так відбува-ється перенесення іонів та дрібних молекул.
Синаптичні з'єднання характерні для нервової тканини, або між нервовою кліти-ною і м'язом, у ділянці якого відбувається передача імпульсу (рис. 6).
| Рис. 6. Міжклітинні контакти: 1-простий контакт; 2-замок; |
| 3-щільний контакт; 4-про- міжний контакт; 5-десмосома; 6-щелеподібний контакт. В1Д ™™™ Д° ^1™™ 3авДЯКИ МЄДіатору. |
Синапс — спеціалізований контакт, що за-безпечує передачу нервового збудження. До його складу входять: ділянка плазмолеми пре-синаптичної мембрани відростка нервової клі-тини, з якої відходить імпульс, ділянка плазмо-леми постсинаптичної мембрани клітини, яка сприймає сигнал. До сигналу входить синап-тична щілина, що розмежовує пресинаптичну та постсинаптичну мембрани і, заповнені не-йромедіатором, синаптичні пухирці. Синапси забезпечують однобічну передачу інформації
Розділ 1
Основи загальної цитологй
Цитоплазма
До складу цитоплазми входять гіалоплазма, органели та включення.
Палоплазма (від гр. пуаііпоз — прозорий) — це прозора рідка скла-дова частина цитоплазми або матрикс, що утворює її внутрішнє серед-овище. При електронній мікроскопії вона має вигляд гомогенної та тон-козернистої речовини з низькою електронною щільністю. Гіалоплазма являє собою складну колоїдну систему і містить різні біополімери: білки, нуклеїнові кислоти, полісахариди тощо. Ця система здатна переходити із рідкого стану в гель і навпаки. Окремі зони гіалоплазми можуть змінюва-ти свій агрегатний стан залежно від умов або функціонального завдання. Деякі молекули білків — тубулінів можуть бути диспергійовані в гіало-плазмі, але в певні моменти вони починають збиратися і утворювати мі-кротрубочки. Таким же чином, у гіалоплазмі можуть виникати і розпада-тися різноманітні фібрилярні, нитчасті комплекси білкових молекул. До ферментів матрикса відносять ферменти гліколізу, метаболізму цукрів, азотистих сполук, амінокислот, ліпідів та інших важливих сполук. У гі-алоплазмі відбувається постійний рух іонів до плазматичної мембрани і від неї, до мітохондрій, ядра, вакуолі. Гіалоплазма є зоною переміщень молекул АТФ. В гіалоплазмі містяться органели та включення.
Органели — постійні мікроскопічні та ультраструктурні утворення, що виконують життєвоважливі функції клітин.
Розрізняють органели загального призначення та спеціальні, а по будові мембранні та немембранні.
Мембранні органели — це складові елементи цитоплазми, відме-жовані від гіалоплазми клітинними мембранами. До них відносяться: мітохондрії, ендоплазматична сітка, лізосоми, пероксісоми, комплекс Гольджі;
Немембранні органели — рибосоми, мікрофіламенти, мікротру-бочки, центросома. Всі названі вище належать до органел загального призначення. Поряд з цим в деяких клітинах існують спеціальні ор-ганели — тонофібрили епітеліальних клітин, міофібрили міоцитів та м'язових волокон, нейрофібрили нервових клітин, війки та джгутики.
Мембранні органели
Мітохондрії. Термін «мітохондрія«(від гр. шііоз - нитка і споп-сігіоп — зернятко) введено Бендою у 1897р. Під світловим мікроскопом мітохондрії мають вигляд круглих зерен та коротких паличок товщи-
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
ною 0.2-2 мкм і довжиною 1-10 мкм. При розгляді під електронним мі-кроскопом кожна мітохондрія має овальну або видовжену форму. У них розрізняють зовнішню гладеньку та внутрішню мембрани, від останньої всередину мітохондрії відходять вирости — кристи. Проміжки між крис-тами заповнені матриксом — електронно-щільною речовиною, у якій ви-являють ниткоподібні утворення товщиною 2-3 нм та гранули товщиною 15-20 нм. Ниткоподібні утворення матрикса являють собою молекули ДНК і РНК, а гранули — мітохондріальні рибосоми. У матриксі і у вну-трішній мембрані містяться білки — ферменти, що забезпечують синтез АТФ шляхом окисного фосфорилювання аденозиндифосфату. Кількість мітохондрій в клітинах значно варіює від поодиноких до сотень. Напри-клад в клітинах печінки вони складають 20% загального об'єму цитоплаз-ми і містять в собі 30-35% загальної кількості білку. Мітохондрії концен-труються у тих місцях цитоплазми, де є потреба в АТФ. Основною функ-цією їх є синтез АТФ, який відбувається в результаті процесів окислення органічних субстратів і фосфорилювання АДФ (рис. 7).
Мітохондрії також беруть участь у регуляції обміну води, депо-нування іонів Са, М§, Р, продукцїї попередників стероїдних гормонів. Мітохондрії здатні рухатися; їх кількість збільшується шляхом поділу, або брунькуванням початкових мітохондрій.
Вони чутливі до різних зовнішніх впливів: голодування, дії рентге-нівських променів, наркотиків тощо.
Ендоплазматична сітка (від гр. епсіо - в середині). її вперше опи-сав К.Портер у 1945 р. Вона являє собою субмікроскопічну органелу, яка утворює внутрішньоцитоплазматичну циркуляційну систему. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених безперервною біомембраною. Розрізняють два типи — гранулярну та агранулярну. Ширина канальців гранулярної сітки від 20 до 1000 нм, з боку гіалоплазми її мембрани покриті рибосомами. Останні беруть участь у синтезі білків на «експорт» і білків-ферментів, а також білків, що нагромаджуються в її цистернах та здатні транспортуватися у ва-куолі комплексу Гольджі, де перетворюються і входять до складу лізо-сом або секреторних гранул. У ряді випадків у канальцях або вакуолях гранулярної ендоплазматичної сітки відбувається модифікація білків, зв'язування їх з цукрами і утворення секреторних гранул. Гранулярна ендоплазматична сітка здатна синтезувати мембранні інтегральні біл-ки, які вмонтовуються в товщу мембран.
Основи загальної цитології
| ',-■-■ ■.'.'■?*:.■ іі*»і*. •• •.. 5Й**'0' |
| ,і:-'У |
| іь. |
| Д |
■*% -■:
••'". ™, 'і.'-'.і, Ч' V ' •'УУс^'
і'. '.'■' ■•' ' ^МЩ^ЗР^З^Щ- *»г'л» ^"*\*Яь
| *&т |

|
ІШ
,■■ #. *
•Г- ч" V !&№%<&&■■ к^-%>&*^ -"
!-•■ А-ІР^Н.'.'^:'^ - -
Палоплазма
Зовнішня мембрана
Внутрішня мембрана
Матрикс Полісоми
Кристи
Рис. 7. Електронна мікрофотографія мітохондрії.
На відміну від гранулярної ендоплазматичної сітки на мембранах агранулярної ендоплазматичної сітки відсутні рибосоми. Діаметр її ка-нальців і міхурців 50-100 нм. Функція гладкої ендоплазматичної сітки пов'язана з метаболізмом ліпідів, синтезом стероїдних гормонів, глі-когену депонуванням іонів кальцію, дезактивацією отруйних речовин, що особливо характерно для гепатоцитів. Мембрана ендонлазматичної сітки безпосередньо контактує з плазмолемою клітини.
Лізосоми ~ мембранні органели. Виявлені і ідентифіковані в кліти-нах ссавців, птахів, земноводних, вони являють собою кулясті струк-тури розміром 0,2-0,4 мкм, містять понад 60 гідролітичних ферментів, здатних до розщеплення біополімерів різної хімічної природи. Із фер-ментів лізосоми містять протеїнази, нуклеази, глюкозидази, фосфата-зи, ліпази. Ферменти локалізуються у лізосомах і утримуються у них за допомогою ліпопротеїнової мембрани, яка обмежує і відокремлює їх вміст від зовнішніх субстратів.
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
Основна фізіологічна функція лізосом — внутрішньоклітинне трав-лення в зв'язку з процесами фагоцитозу, піноцитозу тощо. Залежно від ультраструктурних та функціональних особливостей лізосоми поділя-ють на первинні, їх ферменти знаходяться у неактивному стані; вторинні, або фагосоми (активовані ферменти в них безпосередньо контактують з розщеплюваними біополімерами), а також залишкові тільця, оточені біо-мембраною, нерозщеплені залишки. Поряд з цим лізосоми можуть брати участь у розщепленні власних макромолекулярних комплексів клітин — аутофагоцитозу. В таких лізосомах виявлені фрагменти і навіть цілі ци-топлазматичні структури: мітохондрії, елементи ендоплазматичної сітки, рибосоми, гранули глікогену, тощо. Це може бути доказом їх ролі в про-цесах деградації клітин.
Пероксісоми — субмікроскопічні мембранні органели. Відіграють вирішальну роль у процесах детоксикації клітини. Пероксісоми — тіль-ця розміром 0,3-1,5 мкм, обмежені мембраною, містять гранулярний матрикс, в центрі якого розміщуються кристалоїдноподібні структури із фібрил і трубок. Вважають, що ці органели утворюються на розши-рених кінцях цистерн ендоплазматичної сітки. Вони особливо розпо-всюджені в клітинах печінки та нирок. Ферментні системи пероксісом (каталаза) спрямовані на утилізацію хімічно активного атомарного кисню, а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
Комплекс Гольджі. Перші відомості про цю органелу належать К.Голь-джі, який в 1898р. описав її у складі нервових клітин. Під світловим мі-кроскопом комплекс Гольджі має вигляд сітчастого утворення. Шд елек-тронним мікроскопом він являє собою мембранні структури, що мають вигляд цистерн товщиною до 25 нм, сплющених у центральній частині і розширених на периферії. Окрему ділянку скупчення цих мембран нази-вають діктіосомою. В проміжках між окремими цистернами знаходяться тонкі прошарки гіалоплазми. У комплексі Гольджі завершується процес формування продуктів синтетичної діяльності клітини — її кінцеве глі-козування (рис. 8,9). Комплекс Гольджі бере участь у агрегацп і нагрома-дженні продуктів, синтезованих в ендоплазматичній сітці та їх хімічній трансформації. У цистернах комплексу Гольджі синтезуються поліцу-криди, які комплексуються з білками, що сприяє утворенню мукопро-теїдів, а також виведенню готових секретів за межі клітини. Крім того, комплекс Гольджі забезпечує формування клітинних лізосом. (рис. 10).
Розділ 1 Основи загальної цитології

|
Рис. 8. Комплекс Гольджі в нервових клітинах спинального ганглію х 400:
1-ядро; 2-цитоплазма; 3-фрагмент комплексу Гольджі.

|
| ©О |
Рис. 9. Схема комплексу Гольджі:
1-транспортні міхурці; 2-зріла поверхня мішечків; 3-секреторні міхурці.
 |
| В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія |

Рис. 10. Схема функціонування лізосоми
і внутрішньоклітинного протеолізу за Де Дювом:
1-часточка, що піддається фагоцитозу; 2-мікромолекула; 3-мІкромолекули в стані піноцитозування клітиною; 4-фагосома; 5-ергастоплазма; 6-лізосо-ми; 7-злиття лізосоми і фагосоми; 8-протеоліз частинок і макромолекул(9); 10-екскреція залишків протеолізу; 11-протеоліз у лізосомі з утворенням фа-гоцитозної вакуолі.
Немембранні органели
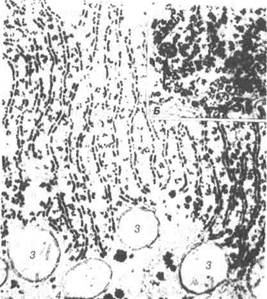
Рибосоми. За розміром рибосоми становлять 25x20x20 нм. До їх складу входять велика і мала субодиниці рибосомної РНК та білок (рис. 11).
Для стабілізації структури рибосом необхідні іони магнію. Кожна із субодиниць побудована з нуклеопротеїдного тяжа, де РНК взаємо-діє з різними білками і утворює тіло рибосоми, у яких амінокислоти сполучаються, тобто синтезуються білкові молекули.
Кілька рибосом, з'єднаних спільною ниткою інформаційної РНК, називають полісомами. Останні пов'язані з мембранами ендоплазма-тичної сітки, синтезують білки для виведення за межі клітини. Сту-пінь інтенсивності синтетичної діяльності вільних рибосом менший, а утворені білки використовуються для внутрішніх потреб клітини.
Мікрофіламенти спостерігаються майже у всіх клітинах. Зокрема в рухових відростках фібробластів та мікроворсинках кишкового епі-
Розділ 1
Основи загальної цитології
телію. Вони мають товщину 5-7 нм, побудовані з скоротливих білків: актину, міозину, тропоміозину та альфа-актину.
9 *?" ГчТт' - \ •- * ' ■ ;* ■ '
А^Г *■*№** V'.
Аі:.- *-і>■-••'■*•,»^*-. І ї
Рис. 11. Електронна мікрофотографія гранулярної