




В організмі постійно відбуваються розщеплення і синтез білків. Обмін білків здійснюється досить інтенсивно. Так, введенням мічених атомів в організм доведено, що вони з високою швидкістю проникають У білки тканин. Однак швидкість оновлення білків різних тканин не однакова, хоч і досить висока. Інтенсивно проходить обмін білків у печінці. В цьому органі він значно іитенсивніший, ніж в інших органах, і поступається лише підшлунковій залозі. Так, в організмі людини білки печінки оновлюються за 10—12 діб, а білки плазми лише за
20—40 діб. Оновлення всіх білків в організмі людини відбувається за 135—155 діб, тобто протягом цього часу всі білкові компоненті! людського організму повністю оновлюються шляхом розщеплення і синтезу.
Основним шляхом розщеплення білків є їх гідролітичне розщеплення до амінокислот. Цей процес протікає не тільки в травному каналі, а й у кожній клітині організму. Гідроліз білків у клітинах різних тканин відбувається в субклітинних утвореннях — лізосомах — під впливом пептидаз, які дістали назву катепсинів). існуют ь катепсини І, II, III, IV, які відрізняються між собою за субстратною специфічністю, і їх дія подібна відповідно до дії пепсину, трипсину, хімотрип-сину і пептидаз. Майже всі катепсици є HS-ферментами.
Нещодавно виявлено ще один шлях розщеплення білків у тканинах за участю нуклеозидтрифосфатів, зокрема АТФ. За цих умов утворюється нуклеотидпептид і поліпептид, укорочений на певну кількість залишків амінокислот:
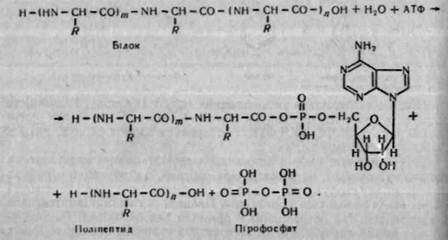
Вважають, що нуклеотидпептиди використовуються для ресинтезу білка у вигляді готових пептидних блоків. Очевидно, такий шлях розщеплення білків має місце при їх перебудові, тобто при перетворенні одних видів білків на інші.
У процесі обміну білків в організмі постійно міститься певна кількість амінокислот. Вони утворюються внаслідок надходження амінокислот з білками їжі, амінокислот, що синтезуються з інших органічних сполук, а також внаслідок розщеплення білків різних органів і тканин. Частина цих амінокислот використовується для утворення білків і ряду інших біологічно важливих речовин, інша частина піддається різним хімічним перетворенням.
ПЕРЕТВОРЕННЯ АМІНОКИСЛОТ
В основі перетворення амінокислот лежать три основні види реакцій: реакції за аміногрупою, за карбоксильною групою і за радикалом амінокислоти. Перші два види реакцій однотипні для всіх L-амі-нокислот. Реакції амінокислот за радикалом досить різноманітні і специфічні майже для кожної амінокислоти.
Перетворення амінокислоти за аміногрупою. До таких реакцій належать два основні види перетворення амінокислот: дезамінування і переамінування.
Дезамінування — процес відщеплення аміногрупи від амінокислоти з утворенням молекули аміаку. Відомо чотири види дезамінування: окислювальне, відновне, гідролітичне і внутрішньомолекулярне.
Під час окислювального дезамінування амінокислоти перетворюються на кетокислоти:

Під час відновного дезамінування амінокислоти перетворюються на насичені жирні кислоти:

Гідролітичне дезамінування пов'язане з перетворенням амінокислот на оксикислоти:

Внутрішньомолекулярне дезамінування характеризується перетворенням амінокислот на ненасичені жирні кислоти:

Зазначені вище реакції каталізують специфічні ферменти. Найбільш поширеним в організмах людини і тварин є окислювальне дезамінування. Розглянемо його механізм на прикладі глутамінової кислоти. Спочатку амінокислота при каталітичній дії глутаматдегідре-генази, коферментом якої є НАД""" або НАДФЬ (для дегідрогеназ інших амінокислот коферментом є ФМН), окислюється до іміноглута-

|
| рової кислоти
ноос-сн2-сн2-сн-соон |
NH,
Глутамінова кислота
-+• НООС—СН2—СН2—С—СООН + НАД • Н, (НАДФ • Н2).
II
Кн
Іміноглутарова кислота
Далі іміноглутарова кислота спонтанно, без участі ферменту гідролі-зує з утворенням а-кетоглутарової кислоти й аміаку:

Слід зазначити, що обидві реакції оборотні. Це свідчить про те, що з а-кетоглутарової кислоти може утворюватись глутамінова кислота.
Іншим видом перетворення амінокислот за аміногрупою є перс нуваїшя (трансамінування) амінокислот. Цей процес перетворення амінокислот дослідили радянські біохіміки О. Є. Браунштейн І М. Г. Кріцман у 1937 р. Вони експериментально встановили, іцо пі час взаємодії глутамінової і піровиноградної кислот можуть утворюватись а-кетоглутарова кислота та аланін без проміжного утворення аміаку ЛДі дані наштовхнули вчених на думку, що аміногрупа з амінокислоти за допомогою ферментативних систем може переноситись на кетокислоти. При цьому амінокислота перетворюсться на кетокислоту, а кетокислота — в амінокислоту. Згідно з зазначеним вище, загальну схему процесу переаміиування можна подати так:
Дослідження  механізму переамінування показало, що в цьому процесі беруть участь ферменти амінотрансферази, коферментом яких Є фосфорний ефір вітаміну BG (піридоксальфосфат). Під час реакції пі; ридоксальфосфат приєднує аміногрупу амінокислоти і перетворюєте J ся на фосфопіридоксамін, після чого віддає аміногрупу кетокислоті і знову перетворюється на піридоксальфосфат. Розглянемо механізм процесу переамінування на прикладі взаємодії глутамінової і піровиноградної кислот. Для зручності піридоксаль-фермент позначим^
механізму переамінування показало, що в цьому процесі беруть участь ферменти амінотрансферази, коферментом яких Є фосфорний ефір вітаміну BG (піридоксальфосфат). Під час реакції пі; ридоксальфосфат приєднує аміногрупу амінокислоти і перетворюєте J ся на фосфопіридоксамін, після чого віддає аміногрупу кетокислоті і знову перетворюється на піридоксальфосфат. Розглянемо механізм процесу переамінування на прикладі взаємодії глутамінової і піровиноградної кислот. Для зручності піридоксаль-фермент позначим^

|
°\
так: >С — фермент.
На першій стадії відбувається взаємодія глутамінової кислоти з піридоксаль-ферментом, в результаті чого утворюється фермент-субстратний комплекс:

Далі відбувається таутомерне перегруповання комплексу, пов'язане з переміщенням атома водню від залишку глутамінової кислоти до піридоксалю, внаслідок чого подвійний зв'язок також переміщується і виникає між атомами азоту і вуглецю глутамінової кислоти:

|
СООН СООН
СН2 СН2
І І
СН2 СНг
СООН СООН
Після таутомерного перегруповання відбувається гідролітичне розщеплення фермент-субстратного комплексу з утворенням а-кето-глутарової кислоти і піридоксамін-ферменту:

|
СООН СООН
І!
C=N—СН2—фермент -f- НОН <± С=0 + H2N—CHa—фермент.
Піридоксамін-фермент, що утворився, взаємодіє з піровиноградною кислотою з утворенням іншого фермент-субстратного комплексу

У цьому комплексі знову відбувається перегруповання:
СН3 СН3
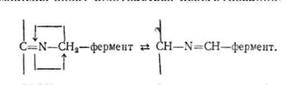
соон соон
Далі відбувається гідроліз комплексу з утворенням амінокислоти (аланіну)  і виділенням піридоксаль-фермепту
і виділенням піридоксаль-фермепту
Отже, у процесі переамінування амінокислота (глутамінова) перетворилась на кетокислоту (а-кетоглутарову), а кетокислота (піровиноградна) — на амінокислоту (аланін). Переамінування амінокислот відбувається у різних органах і тканинах. Воно відіграє важливу роль у процесах синтезу необхідних для організму амінокислот. Кетокислоти, що утворюються внаслідок переамінування, можуть використовуватись для синтезу ліпідів, вуглеводів та інших речовин.
Слід зазначити, що між переамінуванням і окислювальним дезамінуванням існує певний зв'язок. Він полягає в існуванні непрямого дезамінування, тобто в перенесенні аміногрупи а-амінокислоти на a-j кетоглутарову кислоту шляхом трансамінування. При цьому а-аміно-кислота перетворюється на кетокислоту, а глутамінова кислота піддається дезамінуванню  з утворенням а-кетоглутарової кислоти й аміаку!
з утворенням а-кетоглутарової кислоти й аміаку!

Зазначені реакції свідчать про те, що дезамінування вихідної амінокислоти відбувається не безпосередньо, а через трансамінування за участю ос-кетоглутарової кислоти як акцептора аміногрупи.
Перетворення амінокислоти за карбоксильною групою. До таких реакцій належать реакції декарбоксилювання амінокислот з утворенням аміноациладенілатів.
Процес декарбоксилювання амінокислот досить поширений як у тваринних, так і в рослинних організмах. Він пов'язаний з відщепленням від молекули амінокислоти С02 за рахунок карбоксильної групи, в результаті чого утворюються відповідні аміни:

Реакції декарбоксилювання каталізують ферменти декарбоксилази. У вигляді коферменту в їх складі виступає піридоксальфосфат. Винятком є декарбоксилаза гістидину.
Аміни, що утворюються в процесі декарбоксилювання, мають високу біологічну активність, тому їх називають також біогенними амінами. Наприклад, під час декарбоксилювання глутамінової кислоти утворюється у-аміномасляна кислота:

Ця амінокислота відіграє важливу роль у функціональній діяльності центральної нервової системи.
Іншим прикладом є декарбоксилювання гістидину з утворенням гістаміну:

|
—сн2-сн-соон_______ и—j-ov-CHrNH,
Гістамін здатний розширювати судини, знижувати кров'яний тиск, а також активізувати діяльність залоз внутрішньої секреції.

|
Слід зазначити, що аміни виявляють фізіологічну дію при досить малих концентраціях. Нагромадження їх в організмі може викликати порушення ряду біохімічних процесів. Щоб запобігти цьому, в тканинах є активна амінооксидаза, яка каталізує процеси окислення амінів до альдегідів і кислот, частина яких виводиться з організму з сечею, а інша частина піддається перетворенням:
#—CH2-CH2-NH2-{-02 -* R— CH2—CH=NH + Н202;
/?—СН2—CH=NH + НОН -* R— CH,-Cf
ХН
/° /?-CH2-Of + О -* #-СН2-СООН.
NH
Процеси декарбоксилювання більш характерні для мікроорганізмів і протікають дещо обмежено в організмах людини і тварин.
Іншою характерною реакцією амінокислот за карбоксильною групою є утворення аміноациладенілатів. Реакція відбувається за участю специфічного ферменту і АТФ:

Аміноаиилаленілат
Утворення аміноациладенілатів має важливе значення для активації амінокислот, які включаються в процес біосинтезу білка.
Перетворення амінокислот за радикалами. Для амінокислот, крім реакцій за аміно- та карбоксильною групами, властиві хімічні перетворення, пов'язані з реакціями за радикалом.
Характерною особливістю радикалів амінокислот є те, що вони містять різні функціональні групи, рухомі атоми водню тощо. Цим зумовлена активна участь їх в багатьох хімічних реакціях, частина з яких приводить до утворення ряду замінних амінокислот в організмі.
Для прикладу розглянемо реакції окислення окремих амінокислот. Так, під час окислення радикала амінокислоти фенілаланіну утворюється амінокислота тирозин:

Фенілаланін
Реакцію каталізує фермент фенілаланін — гідроксилаза.
Активну участь в окислювально-відновних процесах беруть радикали сірковмісних амінокислот — цистеїну і цистину. Наприклад, під час окислення двох молеул цистеїну утворюється молекула амінокислоти цистину:

Реакція відбувається за участю ферменту цистеїнредуктази, коферментом якої є НАД+.
При повному окисленні сульфгідрильної групи цистеїну утворю
ється цистеїнова кислота, яка під час декарбоксилювання перетворю
ється на таурин: _,

|
О О
II
CH,-SH СН,—S—ОН
......, Окисления '..-_ Декарбоксилювання
LH—Nnt---------------- ► LH—Лпа------------------------------
І І
соон соон
Цистеїн Цистеїнова Таурин
кислота
Важливе значення для організму мають реакції трансметилювання, які здійснюються за участю АТФ. У цих процесах універсальним донором метильної групи є амінокислота метіонін. Спочатку метіонін взаємодіє з АТФ, внаслідок чого утворюється S-аденозилметіонін — активна форма метіоніну:
Цю реакцію каталізує фермент S-аденозилметіонін — синтетаза. Далі метильна група за участю S-метиладенозилметіоніну переноситься на сполуку, яка піддається метилюванню, наприклад на диметила-
І/И?/!


|
міпоетанол. При цьому утворюється холін і виділяється S-аденозил-гомоцистеїн. Реакція каталізується ферментом метилтрансферазою:
Іншим прикладом перетворення амінокислот за радикалом може бути гідролітичне розщеплення аргініну за участю ферменту аргінази. При цьому утворюються амінокислота орнітин і сечовина:

Слід підкреслити, що радикалу кожної амінокислоти властиві певні хімічні перетворення.
Кінцеві продукти обміну амінокислот. Підсумовуючи перетворення амінокислот за аміно- і карбоксильною групами та за радикалами, слід зазначити, що за цих самих умов утворюються аміак, оксид вуглецю (IV), карбонові кислоти, кетокислоти, аміни та інші сполуки. Якщо ці сполуки не використовуються в процесах синтезу, то вони піддаються дальшим перетворенням (крім NH3 і С02). Аміни внаслідок окислювального дезамінування перетворюються на карбонові кислоти, карбонові кислоти і кетокислоти, які утворились внаслідок розщеплення амінокислот, піддаються fJ-окисленню і в трикарбоновому циклі розщеплюються до кінцевих продуктів — Н20 і С02. Отже, кінцевими продуктами розщеплення амінокислот є аміак, вода і оксид вуглецю (IV).
Аміак, що утворюється під час обміну амінокислот у низькоорга-пізованнх організмів, в яких продукти обміну виділяються в порожнину тіла і вимиваються водою, є кінцевим продуктом білкового обміну. Він може виділятись як кінцевий продукт азотистого обміну й у вигляді солей амонію. Такі організми дістали назву амоніотелітичних. До них належать краби, річкові раки, п'явки, каракатиці і деякі інші.
Для організмів людини і високоорганізованих тварин аміак є токсичною речовиною. Особливо шкідливо він впливає на центральну нервову систему, значно порушує процеси тканинного дихання. Тому аміак в процесі утворення відразу ж зв'язується з утворенням неток-сичних для організму сполук.
Одним з важливих методів знешкодження аміаку є взаємодія його з аспарагіновою і глутаміновою кислотами:

При цьому утворюються відповідні аміди. Обидві реакції відбуваються за участю енергії АТФ, їх каталізують ферменти аспарагінсин-тетаза і глут^мінсинтетаза. Аспарагінова і глутамінова кислоти зв'язують аміак, перебуваючи як у вільному стані, так і в складі білків.
Аміак у складі аспарагіну і глутаміну надходить у печінку, де з нього синтезується сечовина, що є кінцевим продуктом обміну білків і амінокислот у більшості хребетних. Такі організми дістали назву уреотелітичних. До них належать ссавці, амфібії, деякі риби, черепахи та інші.
Слід пам'ятати, що утворення сечовини в організмах людини і більшості тварин є основним методом знешкодження аміаку.
На підставі досліджень І. П. Павлова, М. В. Ненцького, С. С. Са-лазкіна, Г. А. Кребса та багатьох інших учених встановлено, що процес синтезу сечовини відбувається в кілька стадій.
На першій стадії утворюється карбамілфосфорна кислота за участю ферменту карбаматкінази, активатором якої, очевидно, є N-ацетил-глутамінова кислота:
"Далі за  участю ферменту орнітинкарбамілтрансферази карбамілфосфорна кислота взаємодіє з орнітином з утворенням цитруліну:
участю ферменту орнітинкарбамілтрансферази карбамілфосфорна кислота взаємодіє з орнітином з утворенням цитруліну:

|
Орнітин Карбамілфосфорна Цитрулін кислота
Під дією ферменту аргінінсукцинатсинтетази за участю АТФ цитрулін взаємодіє з аспарагіновою кислотою з утворенням нестійкої сполуки — аргініпяіітарної кислоти:

Під впливом каталітичної дії ферменту аргінінсукцинатліази аргі-нінянтарна кислота розщеплюється на аргінін та фумарову кислоту: (див. с 370).
Аргінін під впливом ферменту аргінази розкладається на орнітин

|
і сечовину:
NH2—С— NH—(СН2)з—СН—СООН + НаО -*-
II І
NH NH,
Аргінін
NHa—(СН2)з—СН-СООН -f- CO
•NH

Орніті'Н знову може включитись у цикл синтезу аргініну і розщеплення останнього на орнітин і сечовину. Фумарова кислота внаслідок гідратації перетворюється на яблучну, яка при дегідруванні переходить у щавлевооцтову, а остання, приєднуючи аміак, може перетворюватись в аспарагінову.
Слід зауважити, що є ряд фактів, які свідчать про використання сечовини в організмах тварин. Однак це властиве лише для жуйних тварин, які мають багатокамерний шлунок, що містить величезну кількість мікроорганізмів. У рубці карбамід розщеплюється ферментом уреазою, який виділяється мікроорганізмами, до аміаку й оксиду вуглецю (IV). З аміаку і продуктів розщеплення вуглеводів корму мікроорганізми синтезують білок свого тіла, який надходить у сичуг і кишки, де перетравлюється і засвоюється організмом тварини. Саме ця особливість зумовлює широке практичне використання сечовини як часткового замінника білка в раціоні жуйних тварин.
БІОСИНТЕЗ БІЛКІВ
Проблема біосинтезу білка є однією з основних проблем біохімії Вона має важливе теоретичне і практичне значення, тісно пов'язані з найактуальнішими питаннями сучасної біологічної науки: вияснен ням законів спадковості і мінливості, керуванням ростом і розвитко організмів, розкриттям причин виникнення і розробкою методів про філактики та лікування багатьох спадкових захворювань тощо.
Відомо, що в організмах людини і тварин синтезуються мільйон різних білків. Вони відрізняються один від одного, насамперед, хіміч пою природою та послідовністю розташування залишків амінокисло у поліпептидних ланцюгах, тобто первинною структурою/1нформаці про те, яким повинен бути білок, закладена в ДНК у вигляді певно послідовності нуклеотидних залишків у полінуклеотидному ланцюгу
J
Оскільки ДНК знаходиться в ядрі, а біосинтез білка відбувається на рибосомах, то ДНК передає інформацію щодо процесу синтезу білка через ІРНК, яка синтезується на певній ділянці (гені) одного з нуклео-тидних ланцюгів ДНК.
В основі передачі інформації лежить принцип комплементарності. У синтезованій ІРНК послідовність нуклеотидів відповідає послідовності пуклеотидів в одному з полінуклеотидних ланцюгів ДНК. Відмінність полягає лише в тому, що замість тимідинового нуклеотиду в ІРНК міститься уридиновий нуклеотид. Процес копіювання даної інформації з ДНК на ІРНК називається транскрипцією.
Ram ІРНК, діставши інформацію від ДНК, виходить з ядра і переміщується до рибосом. На рибосомах ІРНК реалізує цю інформацію в процесі синтезу білка. Іншими словами, на ІРНК як на матриці відбувається синтез білка, первинна структура якого визначається інформацією, що ІРНК дістала від ДНК.'Процес передачі інформації з ІРНК, яка закодована в певній послідбвцості нуклеотидів в її" молекулі, на процес розміщення залишків амінокислот у білкових молекулах на: зиваєть'ся трансляцією.
Отже, передачу інформації від ДНК на синтез білка можна подати схемою:

Для синтезу білків використовуються активовані форми амінокислот, які перебувають у зв'язаному етапі з відповідними тРНК. Останні переносять їх до місця біосинтезу білка — рибосом. Процес сполучення амінокислот із «своїми» тРНК за участю ферменту аміиоацил-тРНК — синтетази називають рекогніцією (від англ. recognice — пізнавати).
Згідно з сучасними уявленнями, біосинтез білків включає ряд складних біохімічних процесів, в яких беруть участь нуклеїнові кислоти, різні ферментні системи, іони металів тощо. Він поділяється на три основні стадії: транскрипцію, рекогніцію і трансляцію. Стадія транскрипції детально описана в розділі «Нуклеїнові кислоти», стадії рекогиіції і трансляції описані нижче.
Рекогніція
Активація амінокислот відбувається у цитоплазмі за участю високоенергетичної сполуки — АТФ і ферментів — аміноацил-тРНК — синтетаз (аміноацилсинтетаз). Для кожної амінокислоти в клітині є специфічні ферменти. Академік В. О. Енгельгардт назвав ці ферменти кодазами. Для виявл'ення ними максимальної активності необхідні іони магнію. Дещо меншу активуючу дію мають іони інших двовалентних металів, зокрема марганцю, кобальту і кальцію. Реакцію активації амінокислот схематично можна подати так:
.-Vy\

|
0 о
II II
+ НО-Р-0-Р-ОН.
1 I
Он он
'Н
Аміноаииладенілат ОН ОН Пірофосфат
Ця реакція відбувається на поверхні ферменту, що каталізує її, і утворений аміноаииладенілат не переходить у розчин, а залишається в комплексі з ферментом. У молекулі аміноациладенілату залишок амінокислоти сполучається з залишком АМФ макроергічним зв'язком, який посилює реакційну здатність амінокислоти.
На наступному етапі комплекс аміноациладенілату з ферментом взаємодіє з тРНК, специфічною для кожної амінокислоти. При цьому аміпоацильна група з аміноациладенілату переходить до тРНК з утворенням нового комплексу — аміноацил-тРНК і шдаляються АМФ та фермент (див. с 372, 373).
Залишок амінокислоти приєднується до третього атома вуглецю рибози кінцевого нуклеотиду тРНК, макроергічний зв'язок зберігається. Реакція каталізує той самий фермент, що й реакцію активації амінокислоти — аміноацил-тРНК — синтетаза. У молекулі даного ферменту є дві специфічні ділянки, завдяки яким він здатний «впізна-

|
По.їінуклеотидний ланцюг.,,,

другої — «свою»
вати», з однієї сторони, «свою» амінокислоту, а тРНК.
Тепер багато аміноацнлсинтетаз добуто в очищеному стані, а деякі — і в кристалічному. Деякі аміноацилсиитетази (наприклад, вал і нова, лейцинова, ізолейцинова) побудовані з одного пол і пептидного ланцюга, інші — з двох, чотирьох і більшої кількості поліпептидних ланцюгів (наприклад, серинова аміноацилсинтетаза складається з двох субодиниць, метіоніпова — з чотирьох). Молекулярна маса кожної су-бодиниці становить 45 000. Виявлено аміноацилсиитетази, побудовані з двох і більшої кількості різних субодиниць. Наприклад, глі-цинова аміноацилсинтетаза складається з чотирьох субодиниць, дві з яких мають молекулярну масу по 33 000, а дві інші — по 80 000.
Аміноацилсиитетази виявляють високу специфічність. Для кожної з 20 амінокислот, що входять до складу білків, є своя, і притому лише одна аміноацилсинтетаза. Водночас кожна з них «впізнає» всі тРНК, специфічні для однієї амінокислоти.
Трансляція
Трансляція включає три основні етапи — ініціацію, елонгацію і термінацію.
Ініціація синтезу поліпептидного ланцюга — це утворення ініцію-ючого комплексу і формування функціонально активної рибосоми^ Рибосоми — складні рибонуклеопротеїдні комплекси, що складаються з білка і нуклеїнових кислоту У рибосомах нуклеїнові кислоти представлені в основному рибосомальними РНК. Рибосоми здатні сполучатися одна з одною і утворювати пол і рибосоми (полісоми).
(Розміри рибосом характеризують константами седиментації (виражають одиницями Сведберга £)_.) Чим більші розміри мають часточки, тим швидше вони осідають під час ультрацентрифугування і тим вища константа їх седиментації.
Для прокаріот характерні рибосоми, константа седиментації яких становить 705, а молекулярна маса 3 • 10е. В еукаріотів рибосоми дещо більші. Константа їх седиментації досягає 805, а молекулярна маса 4,5 • 10\
Кожна рибосома складається з двох субодиниць — малої і великої. У прокаріот 705 рибосоми побудовані з малої (305) і великої (505) субодиниць. Мала субодшшця містить молекулу 165-РНК і 21 молекулу білків з різною молекулярною масою. 165-РНК виконує в суб-одиниці структурну роль, вона необхідна для контакту рибосом з іРНК. Для контакту з ІРНК у цій самій субодиниці є зона, в якій розміщена спеціальна акцепторна ділянкаЛ(Аср-ділянка) для зв'язування тРНК, яка доставляє активовані амінокислоти.
505 субодиниця містить дві молекули РНК: 235-РНК, 55-РНК і 34 молекули різних білків. 235-РНК виконує структурну роль, а 55-РНК необхідна для взаємодії субодиниці з тРНК. Отже, ІРНК сполучається як з малою, так і з великою субодиницею рибосом,..
На 505 субодиниці рибосоми є дві ділянки — пептидильна, або П-ділянка, і аміноацильна, або А-ділянка; П-ділянка призначена1 для розміщення поліпептидного ланцюга, що росте, а в А-ділянку поступають аміноацил-тРНК.
Рибосоми еукаріот 805 побудовані з малої (405) і великої (605) субодиниць. Мала субодиниця містить молекулу 185-РНК і близько ЗО молекул різних білків. До складу великої субодиниці входять молекули 285-РНК і 55-РНК, а також близько 50 молекул різних білків. На субодиницях рибосом еукаріот також є система зон і ділянок для контакту і зв'язування іРНК, тРНК та ряду інших компонентів, необхідних для синтезу білка.
За певних умов рибосоми можуть розщеплюватись на субодиниці, тобто дисоціювати, а потім знову сполучатися — асоціювати, що має велике значення в процесі синтезу білка. І, Для ініціації синтезу білка крім рибосом і рибонуклеїнових кислот
важливим є наявність формілметіоніл-тРНК (тРНКфЄТ), факторів ініціації (1F-1, 1F-2, 1F-3), ГТФ, іонів магнію тощо. Ініціація синтезу білка найглибше вивчена на прикладі бактерій Е. соїі.
Формілметіонін утворюється після приєднання метіоніну до тРНК за участю 10-формілтетрагідрофолієвої кислоти і специфічного ферменту — трансформілази. Така тРНК за будовою відрізняється від звичайної метіонінової тРНК. Обидві метіонін-тРНК (формілметіо-ніл-тРНК і метіоніл-тРНК) «впізнають» на іРНК триплет (кодон) АУГ. Однак даний триплет кодує формілметіонін лише тоді, коли він є початковим (ініціюючим), тобто коли з нього починаються синтез поліпептидного ланцюга молекули білка. Якщо даний триплет розміщений не в тому місці іРНК, з якого починається синтез білкової молекули, а далі, всередині полінуклеотидного ланцюга іРНК, то він кодує звичайний метіонін.

|
Основна роль формільного залишку в метіоніні полягає в тому, що він, зв'язуючи аміногрупу, зумовлює взаємодію карбоксильної групи першої амінокислоти а аміногрупою наступної амінокислоти, тобто відбувається синтез поліпеп-тидного ланцюга в напрямку NR, -»■ -*- СООН. Після закінчення синтезу пол і пептидного ланцюга від нього може відщеплюватись формільна група або повністю формілметіонін. І Перед початком процесу ініціації рибосома розщеплюється на субодиниці. Ініціація розпочинається з утворення двох комплексів (рис. 64). Перший подвійний комплекс 1F-3—305 утворюється внаслідок взаємодії меншої субодиниці рибосоми 305 з фактором ініціації 1F-3. Другий трикомпонентний 1F-2 • ГТФ • тРНКф" комплекс утворюється внаслідок взаємодії формілметіонін-тРНК з фактором ініціації 1F-2 і ГТФ. Далі ці два комплекси сполучаються в один, який має склад 1F-3 • 305 — IF —
| 3QS Рис. 64. Схема ініціації білкового синтезу (утворення ініціюючого комплексу і формування функціональної рибосоми за А. Уайтом та ін.) |
— 2 • ГТФ • tPHKJ". На наступному етапі цей комплекс за участю фактора 1F-1 взаємодіє з ІРНК, утворюючи ініціюючий комплекс. Останній взаємодіє з 505 субоди-ницею рибосоми, що супроводжується вивільненням факторів ініціації і розщепленням ГТФ на ГДФ і Н3Р04
За цих умов утворюєть-
| ся функціонально активна 705 рибосома (305 • 505-РНК • тРНКфЄТ). Механізм ініціації синтезу поліпептидного ланцюга в еукаріб |
V
. і от
на 805 рибосомах в основному подібний до механізму цього процесу в прокаріот, проте має і специфічні особливості, зокрема число факторів ініціації в еукаріот значно більше, ніж у прокаріот, що, очевидно, вказує на їх регулюючу дію.
^Елонгація (ріст) поліпептидного ланцюга у момент закінчення ініціації формілметіонін-тРНК розміщена в П-ділянці рибосоми, а А-ділянка вільна і може приймати наступну аміноацил-тРНК-

|
| щ о. Основні етапи росту пептидного ланцюга (за А. Уайтом та ін., з окремими змінами і доповненнями автора) |
Перший етап елонгації полягає в доставці аміноацил-тРНК до рибосоми і сполученні її з відповідним кодоном іРНК, розміщеним поряд з початковим (ініціюючим) кодоном АУГ. У цьому процесі беруть участь ГТФ- і фактори елонгації EF — Ти, EF — Ts і EF-G (рис. 65)..Спочатку перший фактор елонгації взаємодіє з ГТФ і аміноацил-тРНК з утворенням потрійного комплексу EF-Tu • ГТФ • амі-ноацил-тРНК. Далі цей комплекс доставляється в А-ділян-ку рибосоми, де й відбувається його дисоціація. Аміноацил-тРНК залишається зв'язаною з рибосомою, а ГТФ гідролізує до ГДФ і відщеплюється від рибосоми у вигляді ГДФ-EF-Tu. Під впливом другого фактору елонгації (EF— Та) цей комплекс розкладається й обидва фактори знову включаються в доставку наступної молекули аміноацил-тРНК в А-ділянку рибосоми. Джерелом енергії служить нова молекула ГТФ.
Па другому етапі елонгації формується пептидний зв'язок в А-ді-лимці, де розміщується доставлена відповідна аміноацил-тРНК. Сюди з П-ділянки переміщується формілметіонін-тРНК і утворюється перший пептидний зв'язок за рахунок етериф і кованої карбоксильної групи формілметіонін-тРНК і аміногрупи, доставленої аміпоацил-тРНК. При цьому утворюється пептидил-тРНК:

Э76

|
CH2-S-CHa
О CHa О H R О
II I II I I II
-* H-C-NH-CH-C-N-CH-C-0 с* тРНК + ОН-тРНК. Пептидил-тРНК
Процес утворення пептидного зв'язку каталізує фермент пептидил-трансфераза.
Вивільнена тРНК утримується в П-ділянці, а пептидил-тРНК — в А-ділянці рибосоми.
На третьому етапі відбувається переміщення (транслокація) пептиди л-тРН К з А-ділянки в П-ділянку. Цей процес проходить за рахунок енергії гідролізу другої молекули ГТФ. Реакцію каталізує білковий фактор EF-G, який часто називають рибосомо-залежною гуано-зинтрифосфатазою.
Під час транслокації з П-ділянки видаляється вільна тРНК, а іРНК переміщується по рибосомі на довжину одного кодону. За цих vmob А-ділянка вивільнюється і може приймати нову амінокислоту з тРНК.
Термінація (закінчення синтезу) поліпептидного ланцюга. Термі-нація поліпептиду розпочинається при появі певних сигналів, якими є триплети УАА, УАГ, УГА/ Ці триплети часто називають беззмістовними, термінаторними або нонсенс-триплетами.
У процесі термінації беруть участь фактори RFi і RF2, які каталізують відщеплення синтезованих поліпептидних ланцюгів від рибосоми. Фактор /?FX реагує на появу УАГ і УАА, фактор RF2 — на УАА і УГА.
В еукаріот термінація проходить за участю лише одногофактора — RF, для дії якого необхідна ГТФ.
Процес термінації відбувається в кілька етапів. Спочатку поліпсп-тидний ланцюг з А-ділянки рибосоми, де утворюється останній пеп-тидний зв'язок, переміщується в П-ділянку. ДалД розривається склад-ноефірний зв'язок між С-кінцевою амінокислотою та її тРНК і білок виходить із рибосоми. Комплекс рибосома • ІРНК • тРНК розкладається, рибосома дисоціює на субодиницк/ Вивільнені рибонуклеїнові кислоти, очевидно, можуть використовуватись у наступних циклах синтезу білка. Даний процес досить складний і повністю ще не вивчений.
Обчислено, що синтез типового білка, побудованого з ~150—ГОЗ залишків амінокислот на рибосомах, триває 1—3 хв. Отже, кожний цикл роботи рибосоми, що забезпечує подовження поліпептидного ланцюга білка на один залишок амінокислоти, триває частки секунди. З'ясування основних етапів синтезу білка є значним досягненням біології на молекулярному рівні. Уже експериментально встановлена генетична роль нуклеїнових кислот, розкрита суть генетичного
коду, основаного на молекулярній структурі ДНК, і цим самим конкретизована природа мутацій — основа еволюції і мінливості живих систем.

