




Ядро — главная часть клетки. Роль ядра: участие в делении клетки, хранение и передача наследственных признаков организма, регуляция процессов жизнедеятельности в клетке, благодаря генетической информации, записанной в молекуле ДНК. В ядре каждой клетки содержится основная наследственная информация, необходимая для развития целого организма со всем разнообразием его свойств и признаков. Именно ядро играет центральную роль в явлениях наследственности.
Андрогенез (мужчина + происхождение) — развитие яйцеклетки с мужским ядром, привнесённым в неё спермием в процессе оплодотворения. Особый случай девственного развития, или партеногенеза; иногда его называют «мужской партеногенез».
Андрогенез наблюдается у отдельных видов животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть женское и мужское ядра не сливаются (Псевдогамия) и в дроблении участвует только мужское ядро.
Астауров экспериментально доказал ведущую роль ядра в наследовании признаков вида и впервые разработал способы направленного получения 100 % особей одного пола на тутовом шелкопряде, заложив тем самым основы теории регуляции пола. Борис Львович первым наблюдал у шелкопряда мутации, индуцированные рентгеновским и гамма-излучением. (http://biografiivsem.ru/astaurov-boris-lvovich).
Хромосомная теория наследственности Т.Г. Моргана. Основные положения. Ограниченность 3 закона Менделя. Кроссинговер и его значение для доказательства линейного расположения хромосом.
Хромосомная теория наследственности - теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности.
Детальная разработка хромосомной теории была произведена Т.Х. Морганом и его учениками (начиная с 1910 г.). Изучая наследование окраски глаз у плодовой мушки дрозофилы, Морган показал, что цвет глаз – признак, сцепленный с полом, и что по характеру его наследования ген, определяющий этот признак, должен находиться в половой хромосоме (Х-хромосоме). Так экспериментально была доказана связь конкретного гена с конкретной хромосомой. В дальнейшем было установлено, что многие признаки наследуются совместно – как один комплекс. Это означало, что контролирующие их гены образуют группы сцепления. Число таких групп сцепления оказалось равным гаплоидному числу хромосом, постоянному для каждого вида организмов.
Затем Морган обнаружил, что сцепленное наследование признаков может нарушаться в результате кроссинговера во время мейоза. На основании детального исследования сцепления генов и кроссинговера (на материале различных мутаций у дрозофилы) Морган и его сотрудники разработали методы определения взаимного положения различных генов на хромосомах и построения генетических карт хромосом. Хромосомная теория нашла подтверждение и дальнейшее развитие в открытии химической природы гена, выяснении строения хромосом и в других достижениях молекулярной генетики.
основные положения хромосомной теории наследственности(1911, Т.Морган)
1. основным материальным носителем наследственности являются хромосомы с локализованными в них генами
2. гены наследственно дискретны, относительно стабильны, но при этом могут мутировать
3. гены расположены в хромосомах линейно в особых участках-локусах на определенном расстоянии друг от друга
4. гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются, как правило, вместе
5. число групп сцепления равно гаплоидному набору хромосом и является постоянным для каждого вида
6. сцепление генов может нарушаться в результате кроссинговера-обмена участками гомологичных хромосом, (это и есть доказательство линейного расположения генов. Кстати, расстояние между соседними генами можно подсчитать. Например, между геном А и В расстояние 3М-морганиды, а между В и С-5 М. Значит, между А и С расстояние будет 8 М)
7.частота кроссинговера прямо пропорциональна расстоянию между генами
Значение теории в том, что она дала объяснение законам Меделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.
Закон независимого наследования (третий закон Менделя) — при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум (и более)парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). Когда скрещивались растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные горошины, наследование каждого из признаков следовало первым двум законам и в потомстве они комбинировались таким образом, как будто их наследование происходило независимо друг от друга. Первое поколение после скрещивания обладало доминантным фенотипом по всем признакам. Во втором поколении наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть 9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами и зелёными горошинами, 1:16 с белыми цветами и зелёными горошинами.
Если в скрещивании будет учитываться большее число генов, чем имеющееся у данного организма число пар гомологичных хромосом, то третий закон Менделя не осуществится. В этом случае вступят в действие другие закономерности, которые будут изложены позднее.
На первый взгляд это ограничение закона независимого сочетания признаков может создать впечатление ограниченности наследственной изменчивости в силу небольшого числа возможных комбинаций гамет. Однако это не так. Рассмотрим для примера возможный размах комбинативной изменчивости, возникающей в силу свободной комбинации гамет у человека. Допустим, что в каждой из 23 пар хромосом человека имеется только по одной паре аллелей. При этом число различных типов гамет выразится величиной 8 388 608, а их возможных комбинаций — 70 368 744 177 664.
Закономерное расщепление при полигибридном скрещивании осуществляется при соблюдении всех условий, о которых шла речь при анализе моногибридного скрещивания. К ним нужно добавить 2 следующих условия: 1) нахождение учитываемых генов в негомологичных хромосомах; 2) равновероятное образование всех сортов гамет на основе случайного расхождения негомологичных хромосом в мейозе.
Кроссинговер (другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус.
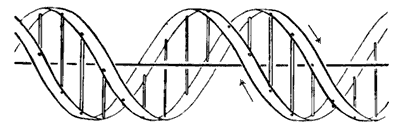
Особенности молекулярного строения ДНК и РНК. Модель структуры ДНК Уотсона-Крика. Правило Э. Чаргаффа. Комплиментарность структуры ДНК. Жесткость молекулярной структуры и ее бесконечная вариабельность. Реакция на выявление ДНК и РНК. Проблема избыточности ДНК.
| Модель структуры ДНК Уотсона и Крика |
Плоскости гетероциклических азотистых оснований расположены внутри двойной спирали перпендикулярно ее общей оси. Расстояние между плоскостями этих гетероциклов по вертикали равно 0,34 нм.
Такая пространственная структура ДНК удерживается множеством водородных связей, образуемых пуриновыми и пиримидиновыми основаниями, направленными внутрь спирали.
Кроме того, стабилизации двойной спирали способствует взаимное перекрывание π-электронных облаков параллельно расположенных ароматических колец нуклеиновых оснований – "стекинг"-взаимодействие (от англ. stack – стопка, штабель).
РНК – это полинуклеотиды, но состоят только из одной цепи, их мол.масса меньше, чем у ДНК. Кроме этого, они отличаются следующим: 1) количество РНК в клетке зависит от возраста, физиологического состояния, органной принадлежности клетки; 2) в мононуклеотидах РНК содержатся рибоза, вместо тимина урацил; 3) для РНК не характерны правила Чаргаффа; 4) в РНК больше минорных оснований, чем в ДНК, при этом в т-РНК количество минорных оснований приближается к 50. Все РНК синтезируются на ДНК, этот процесс называется транскрипцией.
В зависимости от локализации в клетке, функции различают 3 вида РНК: м-РНК (матричная, или информационная), транспортная – т-РНК, рибосомальная – р-РНК.
Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.).
Принцип комплементарности используется в синтезе ДНК. Это строгое соответствие соединения азотистых оснований, соединёнными водородными связями, в котором: А-Т и Г-Ц.
Правила Чаргаффа.
А) Сумма пиримидиновых нуклеотидов равна сумме пуриновых: Пур = Пир
Б) Содержание тимина равно содержанию аденина, а содержание гуанина равно содержанию цитозина: А=Т, Г=Ц.
В) Количество 6-аминогрупп равно количеству 6-кетогрупп: Г+Т = А+Ц.
Г) Отношение А+Т \ Г+Ц видоспецифично.
Биспиральная ДНК весьма жесткая молекула. Это свойство является единым принципом строения ДНК у всех живых организмов. Это говорит о монофилии животного и растительного мира.
Число возможных вариаций ДНК бесконечно. Это объясняет разнообразность и уникальность животного мира.
Реакция на ДНК – реакция Фельгена: реактив Шиффа – фуксин-сернистая кислота (сиренево-фиолетовые глыбки в ядре).
Реакция на РНК – реакция Браше: реактив метиленовый зеленый - пиронин (малиново-красные гранулы в цитоплазме и окрашенные ядрышки).
В молекуле ДНК каждый нуклеотид входит лишь в какой-нибудь один кодон. Поэтому код ДНК неперекрывающийся. Кодоны располагаются друг за другом без перерыва. Так как кодонов возможно 64, то одни и те же аминокислоты могут кодироваться различными триплетами. Такой код называют вырожденным или избыточным.
6)Ген. Эволюция понятия гена. Взгляды Н. Кольцова на биохимическую структуру гена. Требования, предъявляемые к субстрату, ответственному за несение генетической информации.
Ген – участок молекулы ДНК или РНК определяющий последовательность аминокислот в первичной структуре белков. Информационная структура состоящая из ДНК, реже РНК, определяющая синтез молекул РНК одного из типов.
1909 – Йогансен ввел понятие «ген»
1960 – Дж.Симпсон указывает на то, что наследственность осуществляется через гены.
1963 – Ф. Добжанский пишет о том, что единицей наследственности и мутации являются тела молекулярных размеров, названных генами.
В 1927г.- Кольцов сформулировал белковую гипотезу гена. (она просуществовала до 1953 года).
Кольцов рассматривал гены как боковые радикалы огромных белковых молекул (генонем) хромосомы. Гипотеза Кольцова носила умозрительный характер, но она содержала некоторые ценные идеи:
-Ген является химической молекулой, биополимером.
- Ген представляет собой нерегулярный биополимер, несущий в особенностях своего строения специфическую наследственную информацию.
-Гипотеза предусматривала механизм редупликации генетической молекулы посредством матричного синтеза.
Требования, предъявляемые к субстрату наследственности:
-Способность к самовоспроизведению – вещество должно обеспечить преемственность свойств в поколениях
-Уникальность – вещество должно иметь структуру, объясняющую существование миллионов видов и неповторимость.
-Специфичность – структура вещества должна предполагать синтез специфических белков.

