




Пуриндік және пиримидиндік негіздердің ірге тасын екі гетероциклдік негіздер пиримидин мен пуриндер қалайды.

Пурин молекуласы екі конденсияланған сақиналар - пиримидин мен имидазолдан құралған.
 Нуклеин қышқылдарының құрамында үш негізгі пиримидин негіздері- цитозин, урацил және тимин болады.
Нуклеин қышқылдарының құрамында үш негізгі пиримидин негіздері- цитозин, урацил және тимин болады.
 Негізгі пиримидин негіздермен қатар, нуклеин қышқылдарының құрамында минорлы пиримидин негіздері болады: 5-метил- және 5-оксиметилцитозин, дегидроурацил, псевдоурацил, 1-метилурацил, орот қышқылы, 5-карбоксиурацил, 4-тиоурацил және т.б. Тек тРНҚ құрамындағы минорлы негіздердің тізімі 50-ге жақындап қалады. Минорлы негіздердің үлесіне 10 % -ке дейінгі тРНҚ дағы барлық нуклеотидтердің үлесі қамтиды, бұл физиологиялық тұрғыдан өте маңызды (РНҚ молекуласын гидролиздік ферменттердің әрекетінен қорғайды). Кейбір минорлы пиримидин негіздерінің қатарларының құрылымдық формулалары –нуклеозидтер – азоттық қосылыстардың көмірсулармен байланысы түрінде көрсетілген.
Негізгі пиримидин негіздермен қатар, нуклеин қышқылдарының құрамында минорлы пиримидин негіздері болады: 5-метил- және 5-оксиметилцитозин, дегидроурацил, псевдоурацил, 1-метилурацил, орот қышқылы, 5-карбоксиурацил, 4-тиоурацил және т.б. Тек тРНҚ құрамындағы минорлы негіздердің тізімі 50-ге жақындап қалады. Минорлы негіздердің үлесіне 10 % -ке дейінгі тРНҚ дағы барлық нуклеотидтердің үлесі қамтиды, бұл физиологиялық тұрғыдан өте маңызды (РНҚ молекуласын гидролиздік ферменттердің әрекетінен қорғайды). Кейбір минорлы пиримидин негіздерінің қатарларының құрылымдық формулалары –нуклеозидтер – азоттық қосылыстардың көмірсулармен байланысы түрінде көрсетілген.
Нуклеин қышқылдарын гидролиз жүргізгенде алынатын ертінділерде құрамында болатын екі пурин негіздері келесідегідей құрылыста болады:
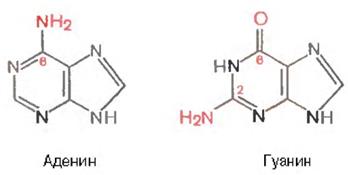
ДНҚ мен РНҚ құрамындағы пуриндік қатарлардың минорлық негіздеріне инозин, N6-метиладенозин, N2-метилгуанозин, ксантин, гипоксантин, 7-метилгуанозин және т.б. жатады. 
Бос азоттық негіздердің басты қасиеттернің бірі,(құрамында оттегі топтары бар) олардың екі таутомерлік формада кездесуі, соның ішінде ортаның рН-на байланысты лактим- және лактамды формаларда болуы: ортаның рН 7,0-ге тең болғанда лактамды формада, рН-тың мөлшері төмендегенде лактимді формада кездеседі. Таутомерлі өзгеруді урацилдің мысалында былай бейнелеуге болады.
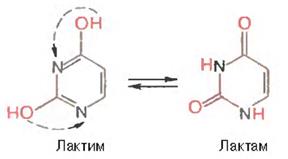
Табиғи жағдайлардағы нуклеин қышқылдарының құрамындағы пуриндер мен пиримидиндердің оттекті негіздері лактамды формада кездеседі.
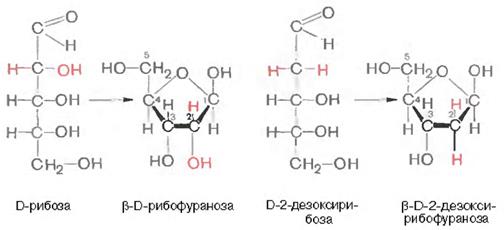
- Көмірсулы компоненттер. ДНҚ молекуласында көмірсу дезоксирибоза түрінде, ал РНҚ молекуласында –рибоза түрінде кездеседі, осы себептен олардың аттары-дезоксрибонуклейн (ДНҚ) және рибонуклейн (РНҚ) қышқылдары..
Көмірсулар (рибоза мен дезоксирибоза) ДНҚ мен РНҚ молекулаларында (рибоза и дезоксирибоза) β-D-рибофуралозды форма түрінде кездеседі:
Кейбір фактор ДНҚ құрамында глюкоза табылған, ол гликозидік байланыс арқылы 5-оксиметилцитозинмен байланысқан.
4. Нуклеозидтер. Нуклеотидтер. ДНҚ-ның бірінші реттік құрылымы. Нуклеин қышқылдарының құрылымдық бірлігін мономерлі молекулалар-мононуклеотидтер құрайды. Демек, нуклеин қышқылдары полинуклеотидтер болып табылады. Бұл мононуклеотидтердің полимерлік өнімдері, олардың ДНҚ мен РНҚ тізбектеріндегі саны мен жүйелілігі матрица молекуласында жазылған бағдарлама арқылы қатаң қадағаланады. Мононуклеотидтер нуклеазаның қатысуымен ДНҚ және РНҚ гидролиздеу барысында пайда болады, ол арнайы компоненттерден тұрады: азоттық негіздер, көмірсу және фосфор қышқылынан тұрады. Бұл «үштіктің» көмірсу ортасында орын тебеді. Азоттық (кез-келген) негіздің көмірсумен (рибоза немесе дезоксирибозамен) байланысы нуклеозид деп аталады, ол мононуклеотидттерден негізбен әрекеттестіргенде, немесе арнайы фермент- нуклеотидаза ферментімен гидролиздеп фосфор қышқылын бөлу барысында түзіледі.
Нуклеозидтердің құрамында пурин немесе пиримидин негіздері болады, олар көмірсумен байланысты N-гликозид арқылы орнатылған. Нуклеин қышқылдарының құрамында тек қана β-нуклеозидттер болады. Мысал ретінде екі мононуклеотидті келтіруге болады: аденозин-5'-монАМФ) және цитидил-5-монофосфор қышқылы (ЦМФ):

ДНК немесе РНК нуклеин қышқылдарының типтеріне сәйкес R 2' көмірсуда Н- немесе ОН-топтары орналасқан. N-гликозид байланыстарының түзілуіне пурин нулеотидтерінде пуриннің 9-N мен пентозаның 1'-С атомдары қатысады, ал пиримидин нуклеотидтерінде пиримидиннің 1-N мен пентозаның 1' –С атомдары қатысады.
Рибоза мен дезокирибозаның құрамындағы көміртегі атомдарын пурин мен пиримидин негіздерінің құрамындағы көміртегі атомдарынан ажырату үшін алғашқы рет «штрих» символдары қолданылады: мысалы, көміртегінің 3-ші және 5-ші атомдары С-3' және С-5' немесе көбінесе 3' және 5' таңбаланады.
ДНҚ мен РНҚ-ға ферменттік гидролиз өткізгеннен кейін алынатын өнімдердің ішінде 5'-нуклеозид монофосфаттармен қатар, 3'-нуклеозид- монофосфаттар да түзілетіндігін осы жерде атауға болады. Фосфордың орны көршілес нуклеотидттердің арасындағы фосфодиэфирлік байланыстың үзілген жерінде болғанын байқауға болады, яғни нуклеотидттер бір-бірімен әр пентозадағы 3' және 5' көміртегі атомдарының арасындағы байланыс фосфор қышқылының қалдықтары арқылы жүретіндігін көрсетеді.
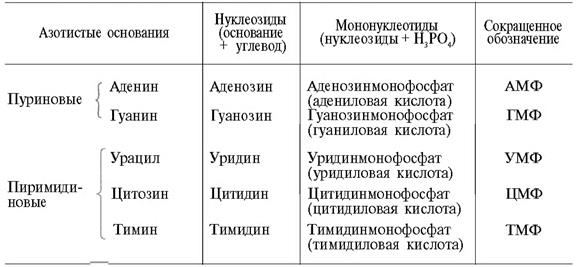
1 кестеде нуклеотидттер мен нуклеозидттердің құрамы аттары (тривиальдық атауы) берілген, сонымен қатар қысқартылған атаулары (РНҚ- да рибонуклеотидтер, ал ДНҚ-да дезоксирибонуклеотидттер).
1 Кесте. Нуклеозидттер мен мононуклеотидттердің құрамы.
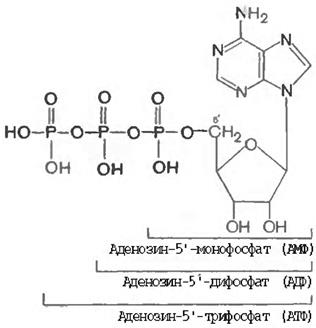
Мононуклеотидттер және олардың туындысы, сонымен бірге динуклеотидттер жасушаларда бос күйінде кездеседі және зат алмасуда маңызды роль атқарады. Сонымен, қатар көптеген коферменттердің құрамына нуклеотидттер кіреді, оның қатарына оксиредуктаза коферментін кіргізуге болады. Мононуклеотидттер енді бір фосфаттың қалдығын қосып алып, фосфоангидридтік байланыс түзіп (пирофосфаттың құрамындағы байланысқа ұқсас) нуклеозиддифосфаттарға айналады (өздеріне тиісті, олар қысқаша (АДФ, ГДФ, УДФ, ЦДФ және ТДФ) деп аталады. Соңғылары, тағы бір фосфат қалдығын қосып алып нуклеозидтрифосфаттарға айналады (олар өздеріне сәйкес АТФ, ГТФ, УТФ, ЦТФ және ТТФ) деп аталады.
Бұл жерде, жасушада бос нуклеозидтрифосфаттар ДНҚ мен РНҚ ферменттік синтезделуі барысында негіз болып келеді. Бірақта, клеткада бос күйіндегі, белок синтездеуге қатыспайтын, бірақта маңызды тіршілік қызмет атқаратын табиғи нуклеозидтрифосфаттар кездеседі. Сонымен, бірге нуклеозидтрифосфаттардың оның ішінде АТФ маңызды функцияларының бірі барлық тірі ағзалардың биоэнергетикасына қатысу. Аденозинди- және аденозинтрифосфаттар молекулаларының түзілу нұсқасын (пурин ядросы мен рибоза сақинасындағы кейбір сутегі және көміртегі атомдары жазылмаған):
Нуклеотидттердің фосфорлы эфирлерлерінің екі типі кездесетіндігін бұл жерде атауға болады, фосфат кейде бір нуклеотидтегі пентоза қалдығының 2- оттегі атомын біріктіреді және кейде фосфат көпіршесі екі әртүрлі мононуклеотидттерді біріктіреді. Бірінші типке мысалы ретінде 2’,3’— және 3’,5’- циклді нуклеотидттерді жатқызуға болады, яғни екі мүмкін болатын қосылыстар класстарында С-2’ и С-3’-көміртегідегі немесе С-3’ и С-5’ -көміртегідегі оттегі атомдары циклды құрылым түзуге қатысады.
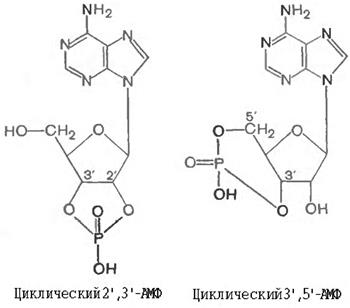
Бұл қосылыстардың біріншісі 2’,3’-АМФ рибонуклеин қышқылдарының ыдырауының аралық өнімі ретінде түзіледі, ал циклді 3’,5’-АМФ (цАМФ) нағыз кездесетін рибонуклеотид болып есептеледі (аденилатциклаза ферментінің катализдеу реакциясынан АТФ –дан түзіледі. цАМФ ерекше, белгілі функциялар атқаруға қабілетті және зат алмасу процессін реттеуде жоғары биологиялық белсенділік көрсетеді, мұнда ол жануарлар жасушасында клетка ішіндегі сигналдардың медиаторы ретінде роль атқарады.
Тап осындай функцияны цГМФ, УДФ, ЦТФ туындылары атқарады және кофакторлар мен коферменттер құрамындағы нуклеотидтер атқарады.
Бір фосфат тобының екі әртүрлі рибонуклеотидті байланыстырған мысалы, ретінде гуанозилтимидинфосфаттың құрлысын келтіруге болады:
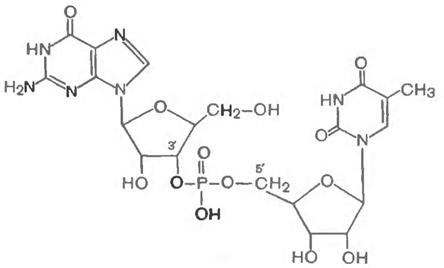
Бұл қосылыстар табиғатта кездеспеген мен, бірақта табиғиғи нуклеин қышқылдарына тән байланыстар осы қосылыстардың құрамынан табылған. Бірінші нуклеотидттің рибозасындағы 3'-С-тегі, фосфодиэфирлік байланыс арқылы екінші нуклеотидттің рибозасындағы 5' –С -тегімен байланысқан.
Медициналық практикада, оның ішінде онкологияда азоттық негіздердің туындылары мен қатар нуклеозидттер мен нуклеотидттер кеңінен қолданылады. Негіздердің немесе көміртегінің құрамында аздап модификацияға ұшыраған бұл аналогты қосылыстар клетка құрамына кіріп алып айырықша цитотоксикалық эффект көрсетеді. Кеңінен таралған дәрілік препараттарға –пуриндік және пиримидиндік негіздердің аналогтары (нуклеотидттерге сәйкес келетін) 5-фторурацил, 6-тио- және 6-меркаптопурин, 8-азагуанин, 6-азауридин және 6-азацитидин және сонымен қатар дезоксиуридиннің 5-иодтты туындылары жатады.
Нуклеозидтер мен нуклеотидтердің қысқаша атауы мен қатар нуклеозидтерді әріптермен аденозин (АМФ) А-мен, гуанозин (ГМФ) Г-мен,цитидин (ЦМФ) Ц-мен уридин (УМФ) У-мен, тимидин (ТМФ) Т-мен таңбалау қолданылады:
Осы символдарды пайдалана отырып жоғарыда көрсетілген дирибонуклеозидмонофосфат Г-Т деп таңбаланады. Құрылысы және қасиеттері жағынан Г-Т және Т-Г бір – бірінен үлкен айырмашылықтары болады (осындай дипептид жағдайында да байқалады).
5. ДНК-ның бірінші реттік құрылысы. Нуклеинқышқылдарының бірінші реттік құрылысы деп ДНҚ мен РНҚ-ның полинуклеотидттік тізбектеріндегі мононуклеотидттердің орналасу ретін айтады. Мұндай тізбек 3',5'-фосфодиэфир байланысының көмегімен қалыптасады. Нуклеин қышқылдарының молекулалық массасы (2 ·104 - 1010 - 1011 аралығында жататындықтан барлық белгілі РНҚ-лардың, оның ішінде ДНҚ бірінші реттік құрылысын анықтау өте күрделі. Бұлай болғанмен, барлық нуклеин қышқылдарында (дәлірек, дара тізбекті нуклеин қышқылдарында) көршілес нуклеотидттердің арасында байланыстың тек бір ғана типі - 3',5' –фосфодиэфир байланысы болады. Бұл құрылымның негізін төмендегідей күйде көрсетуге болады:
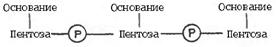
Нуклеотидттер арасындағы байланысты қалыптастыруға 3'- және 5’-көміртегі қалдықтарының гидроксил топтары қатысады.
Қазіргі уақытта барлық тРНҚ, 5S рРНҚ молекулаларының кейбір түрлері, E.coli-дің 16S рРНҚ-ның, вирустық РНҚ-лардың бірінші реттік құрылыстары анықталды, олардың құрамында жүздеген немесе мыңдаған нуклеотидттер қалдықтары болды. Барлық клеткалық РНҚ незінен тек бір тізбектен тұрған. РНҚ молекуласының бір ұшында бос монофосфорлы эфир орналасқан, ол 5´-ұшы, оның қарама-қарсы ұшында мұндай фосфат болмайды, ол жерде бос 2´- және 3´-гидроксил топтары орын тепкен. Егер РНҚ молекуласын сілтілік гидролизге ұшыратсақ, онда полинуклеотидттік тізбектің ең шеткі нуклеотиді ретінде 5´-ұшында бос фосфаты болатын ЦМФ, және 3´-ұшында бос нуклеозид түріндегі аденозин орналасқан.
Қазіргі уақытта ДНҚ әр түрлі молекулаларының бірінші реттік құрылысы анықталған. 30 жыл бұрын адамның митохондриялық ДНҚ нуклеотидттік қатарлары толық анықталған (16569 нуклеотидттер жұбы). Кейбір вирустар мен плазмидттердің ДНҚ нуклеотидттік қатарлары белгілі. Екі прокариотттық организмдердің (Haemophylus influenzae және Mycoplasma genitalum) нуклеотидттік қатарларын анықтау (және алғашқы эукариоттық организм-ашытқы геномының толық оқылуы туралы хабарлар) болды. Мұндай зерттеулер Е.coli және нематод (Caenorhabditis elegans) геномдарымен жүргізілген. Дүние жүзі зерттеущілері адам геномын толық зерттеуге бағытталған жұмыстарын екпінді орындауда.
Әртүрлі ДНҚ молекулаларын секвендеу (нуклеотидттік қатарларын анықтау) қорытындылары компьютерлік банкқа жинақталады, және бұл мағыұлыматтарды дүние жүзілік компьютерлік жүйелерді қолданушылар оңай пайдалана алады.
Соңғы уақытта ДНҚ-ның бірінші реттік құрлысын (дәл айтқанда, оның жекелеген бөліктерін) кейбір жанама көрсеткіштер, мысалы ДНҚ молекуласындағы кейбір нуклеотид аймақтарының жұптасу дәрежесінің көмегімен (мұндай тәсілмен анықтау барысында изоплиттер деп аталатын нуклеотидтер қатарының жекеленген фракцияларының құрылысы мен олардың сандары анықталады, және сонымен қатар ДНҚ-ның реассациялану реакциясының кинетикасы арқылы анықталған (бұл тәсілмен нуклеотидтердің қайталанатын қатарларының саны анықталады). ДНҚ-ның бірінші реттік құрылысын ДНҚ-дағы минорлық негіздердің таралу дәрежесі бойынша (олардың ДНҚ орналасуының белгілі заңдылықтары бар) және ДНҚ-да осы паллиндромдардың табылуы мен олардың құрылыстарының анықталуы(«кері тепкен қатарлар» немесе құбылмалылар) негізінен олар рестрикциялар аймағында орналасады. ДНҚ-ның бірінші реттік құрылысын анықтауда ғалымдар физикалық, химиялық (гендердің синтезі), генетикалық және басқа тәсілдерге, табиғи көздерден кейбір гендерді (немесе олардың фрагменттерін бөлу), және кері транскрипция ферментінің көмегімен мРНҚ-дан түрлі гендерді синтездеу тәсілдеріне үміт артады. ДНҚ–ның бірінші реттік структурасын анықтау үшін жақын арада экспресс метод қолданылған, ол екі ДНҚ- полимераза (E.coli мен T4 бактериофагынан бөлінген) ферментін пайдалану арқылы іске асады. Бірақта жоғарыда айтылған жағдайлардың бәрінде ДНҚ-ның азғантай аймағының құрылысы анықталған, сондықтан адам геномының бірінші реттік структурасын толық анықтау өз шешімін күтуде

