




1. «Молекулалық биология» пәнінің анықтамасы. Молекулалық биологияның тіршілік жүйесі туралы ғылымдар ішіндегі ролі.
2. Белоктар мен нуклеин қышқылдары молекулалық биологияның негізгі зерттеу объектілері. Нуклеин қышқылдары мен белоктардың тірі материяның қасиеті мен тіршілік әрекетін қалыптастырудағы ролі.
3. Молекулалық биологияның орталық догмасы
4. Молекулалық биологияның қысқаша тарихы және даму этаптары
5. ХХ ғасырдың аяғында молекулалық биологияның жетістіктері. Қазақстанда молекулалық биологияның дамуы
1. Молекулалық биология» пәнінің анықтамасы. Молекулалық биологияның тіршілік жүйесі туралы ғылымдар ішіндегі ролі.
Молекулалық биология ХХ ғасырдың екінші жартысында пайда болды. Бұл ғылымның дамуын У.Эстбюрдің атымен байланыстырады, 1939 жылы ол өзін молекулалық биологпын деп атаған.
Ол екі жылдан кейін алғашқы рет ДНҚ рентгенограммасын алған, сол себепті «ең басты молекуланың«дәл құрылымын зерттеудің бастамасын жасады. Бірінші рет молекулалық биология деген ұғымды атаған У. Уивер. Ол Рокфеллер фондысының жаратылыстану бөлімін басқарған. Ол 1938 жылы былай жазған еді: химия мен физиканың биологиямен шектесетін шекаралас аймақтарында біртіндеп жаңа ғылым саласы молекулалық биология дамып отыр, ол жапқышпен көмкеріліп тұрғандай, әлі белгісіз болған тірі клетканың басты қасиеттерін аша бастады. Осындай жағдайда қазіргі биологияның жаңа бағыты туралы алғашқы постулат пайда болды, ол биологтардың, химиктердің және физиктердің тірі табиғат объектерін зертеудегі ізденістерін біріктіріп алға қарай дамытуда үлкен үлес қосты.
Оның ары қарай дамуына биохимиядағы, цитологиядағы және генетикадағы шешілмей тұрған проблемалар себеп болды. Осындай, ХХ ғасырдың ортасындағы қалыптасқан биологиялық ғылымдар молекулалық биологияның дамуына жол ашып, өздерінің алдына тіршілікті молекулалық деңгейде зерттеуге мақсат етіп қойды.
Молекулалық- биологиялық зерттеулердің орталық мәселелері- тұқым қуалаушылықтың материалдық негіздері, гендер табиғаты және ұрпақтан-ұрпаққа тұқым қуалаушылық белгілері мен олардың берілу механизмдері.
Жаңа формация бағытындағы генетиктер (Т.Морганның, Н.К. Кольцовтың, Н.В.Тимофеев-Ресовскийдің және басқалардың) және АҚШ -тан Европаға 30 жылдардың аяғындағы эмиграцияға кеткен физик-теоретиктер мен экспериментаторлар М. Дельбрюктоң басшылығымен онда «фагтар зерттеу тобын» құрған. Олар вирустар мен бактериофагтардың мутагенезы мен молекулалық құрлысын зертеуді бастаған. Кейін бұл жұмыстар бұрынғы ҚССРО елінде Б.Ф. Поглазовтың, Н.А.Киселевтың және басқа ғалымдардың зерттеулерінің арқасында тереңірек дамыған. Бұдан сәл бұрынырақ В.А. Энгельгардт М.Н. Любимова бұлшық еттің созылып-жиырлу механизмін ашса, А.Н. Белозерский өсімдіктен бірінші рет ДНҚ –ны өсімдіктен бөліп алған, бұлар да молекулалық биолгияның іргелі ғылыми бағыттарының бірі. Бұл әйгілі ғалымдардың есімдері кейіннен ірі ғылыми-зерттеу орталықтарына берілді: В.А.Энгельгардт атындағы ҚСРО -ғылым академиясының Молекулалық биология бойынша зерттеу институты, М.Ломоносов атындағы Москва мемлекеттік университеті жанындағы А.Н.Белозерский атындағы физикалық-химия зерттеу институты.
ХХ - ғасырдың 50-ші жылдарының басында биохимияда белоктар мен нуклеин қышқылдарының қарапайым құрылысы туралы іргелі жаңалықтар алынып, онда полипептидтік тізбектерінің ұйымдасу дәрежесі, оның ішінде нуклеотидтердің құрылысы мен олардың ДНҚ мен РНҚ молекулаларындағы кездесетін сандық мөлшерлері заңдылығы ашылды (Э.Чаргафф). Бұл көрсетілген жұмыстар, сонымен қатар Англияда, Розалинд Франклин мен Морис Уилкинсон жүргізген рентгенструктуралық анализдің көмегімен жүргізілген ДНҚ структурасының биофизикалық зерттеулері «фагты зерттеу тобының «шәкірті Джеймс Уотсон мен англиялық физик Френсис Крикка гендердің молекулалық табиғаты мен олардың ДНҚ құрамындағы өзін-өзі көбейте алу (репликация) механизмдерін ашуға көмек болды.
2. Белоктар мен нуклеин қышқылдары молекулалық биологияның негізгі зерттеу объектілері
Организмдердің структуралық-функциональдық бірлігінде тіршіліктің негізін құрайтын белоктар (белоктық денелер) басқа органикалық қосылыстар алмастыра алмайтын заттар қатарына жатады.
Белоктар –бұл жоғары молекулалы азотты органикалық қосылыстар, олардың молекуласы аминқышқылдары қалдықтарынан құралған. «Протейн» атауы (грекше -protos –бірінші, маңызды) бұл классқа жататын заттардың басты биологиялық маңыздылығын айқындай түседі. отандық әдебиеттерде қабылданған «белоктар» және «белоктық заттар» деген терминдер жануарлар мен өсімдіктер ұлпасында тауық жұмыртқасының белогына ұқсас заттардың болуына байланысты қойылған атаулар. Біздің уақытымызда, тұқым қуалаушылық ақпарат кез-келген тірі организмдер клеткасының ДНҚ молекуласында орналасқандығы белгілі болғандықтан, тек белоктардың генетикалық ақпаратты іске асыратын құрал екендігі болып табылатынына күмән жоқ.. Белоксыз, дәл айтқанда ферментсіз ДНҚ-да репликация өтпей, яғни өздігінен екі еселене алмайды, сол себепті генетикалық ақпаратты тасымалдауға қабілетсіз.
Тірі табиғаттың, өлі табиғаттан ерекшелендіретін бірнеше қасиеттері болады, ол қасиеттердің бәрі белоктардың қызметіне байланысты. Негізінен, барлық тірі ағзалардың белоктары құрылысы жағынан көптүрлі болып, құрылысы кеңістік және уақыт аралығында жоғары дәрежеде белгілі ретке келтірілген. Тірі ағзалардың ерекше қасиеттерінің бірі өзі сияқты ағзаларды өндіруі де, белоктардың қызметіне байланысты. Бұлшық еттердің жиырлып жазылуы, қозғалысқа келтірілуі тіршілік жүйесінің міндетті атрибуты-бұлшық еттер аппаратының белоктық құрылымдарының жұмысына байланысты. Сайып келгенде, зат алмасу жүрмесе, тірі ағзаның құрамдас бөліктерінің тұрақты жаңарып отыруы жүрмесе, яғни анаболизм мен катаболизм үдерістері өтпесе (мұндай тіршілік нысанына кері таңқаларлық бірлігі) тіршілікті көзге елестету мүмкін емес, олардың негізін катализдік активті белоктар-ферменттердің қызметтері құрайды.
Сонымен, белоктар (белоктық заттар) тірі ағзалардың тірішілік негіздері мен құрылымдарының негіздерін құрайды. Молекулалық биологияның негізін қалаушы Ф.Криктің сөзі бойынша белоктар әр түрлі қызметтерді таңқаларлық түрде жеңілділікпен және әдемілікпен жасай алады. Есептелгендей, табиғатта 1010-10 12 әртүрлі белок кездеседі, олар құрылысының күрделілігі жағынан түрліше ұйымдасқан, вирустардан басталып адамға дейінгі, 10 60 тіршілік формаларының пайда болуын қамтамасыз етеді.
Нуклеин қышқылдары басқа полимерлік негіздерге тән емес маңызды биологиялық функцияларды атқарады. Дәлірек айтқанда, олар тіклей тұқым қуалаушылық ақпаратты сақтау мен ұрпақтан-ұрпаққа тасымалдауды және ақпараттың тасымалдау механизмін жасушадағы барлық белоктарының программасын жасақтау арқылы іске асырады. Бұдан басқа, нуклеин қышқылдар құрамына кіретін құрылымдық бөліктері кофакторлар (А- коэнзим, уридин-дифосфатглюкоза және т.б.), аллостерикалық эффекторлар ретінде қызметін атқара алады және кофферменттердің (никотинамидадениндинуклеотид, флавинадениндинуклеотид және басқалар) құрамына кіріп зат алмасу үдерісіне тікелей қатысып, сонымен қатар энергияның жинақталуы, тасымалдануы және бір күйден екінші күйге ауысуын қамтамасыз етеді. Олар жасуша ішінде пайда болған маңызды сигналдарды тасымалдайтын, екінші реттік делдарлардың (мессенджерлердің) – циклді мононуклеотидтердің (цАМФ және цГМФ) –ізашары (предшественник) ретінде қызмет атқара алады.
3. Молекулалық биологияның орталық догмасы
ДНҚ структурасының биофизикалық Дж. Уотсонның, Ф. Криктің, Х-Г. Корананың, А. Корнбергтің және ХХ- ғасырдың 60- жылдарының ортасындағы Нобель силығына иегер болған ғалымдар молекулалық генетиканың негізгі постулатын ашты, ол клеткадағы генетикалық информацияның берілу механизмдерінің іске асуын түсіндіреді.
ДНК → РНК → Белок
Одан кейін ДНК өзін-өзі ұлғайту (репликация) механизмі, транскрипция (РНК биосинтезі) және трансляция (белок биосинтезі) зерттелді. Осыған параллелді бағытта бұл процестердің клетка ішінде орын тебуі зертелді, бұл клетка ішілік компоненттердің (ядроның, митохондрияның, рибосоманың және басқалардың маңыздылығын түснуге жол ашты және осы мағұлыматтар Дж. Уотсонға 1968 жылы молекулалық биологияға мынандай анықтама беруге негіз болды:» Молекулярлық биология биологиялық макромолекуланың структурасын және негізгі клеткалық компоненттердің функциялармен байланысын зерттейді, және сонымен қатар клеткалардың өздігінінен реттеу механизмдерінің негізгі принциптерін және барлық процестердің бірлігін реттеп, тіршілікті қалыптастыратын барлық процестерлің өз-ара келіслуін болғызып отырады.
Кейінен молекулалық генетиканың орталық догмасы кері транскрипциясы болатындығы туралы жаңалық ашылғаннан кейін төмендегідей түрде өзгеріске ұшырады.

4. Молекулалық биологияның қысқаша тарихы және даму этаптары
1869 жылы – Ф. Мишер (F. Miesher) алғаш рет лейкоциттер мен лосось тұқымынан ДНҚ бөліп шығарды.
1935 жылы – А.Н. Белозерский өсімдіктерден ДНҚ бөліп алды.
1939 жылы – В.А. Энгельгардт миозинның АТР-дық белсенділігін ашты.
1940 жылы – У. Эстбюри (W. Astbury) ДНҚ-ның бірінші рентгенограммасын алды.
1944 жылы – О.Т. Эвери (O.T. Avery) генетикалық ақпаратты тасымалдаушы ДНҚ (бұрынғы жорамалдағандағыдай белок емес) екендігін жариялады.
1951 жылы – Л. Полинг және Р. Кюри (L. Pauling, R. Corey) полипептидтік белок тізбектерінде аминқышқылдық қалдықтардың шоғырлануының негізгі түрлерінің бар екендігін дәлелдеді.
1953 жылы – Дж. Уотсон мен Ф. Крик (J. Watson, F. Crick) Р.Франклин мен М.Уилкинс (R. Franklin, M. Wilkins) алған рентгенограмма негізінде ДНҚ-ның қос спиральдік үлгісін жасады.
1953 жылы – Ф. Сангер (F. Sanger) өгіздің алғашқы инсулиндік құрамын бағалады.
1956 жылы – А. Корнберг (A. Kornberg) ДНҚ-полимеразасын ашты.
1957 жылы – А.Н. Белозерский мен А.С. Спирин мРНҚ-ның бар екендігі туралы жорамалдады.
1960 жылы – Дж. Кедрью (J. Kendrew) кашалот миоглобинінің үшөлшемдік құрылымын, ал М. Перутц (M. Perutz) – гемоглобин құрылымын алғаш рет хаттады.
1960 жылы – бір мезгілде бірнеше зертханаларда транскрипция ферменті – РНҚ-полимераза ашылды.
1961 жылы – Ф.Жакоб пен Дж. Моно (F. Jacob, J. Monod) оперон үлгісін жасады.
1965-1967 жылдары – Р. Холли (R. Holley) аланиндік тРНҚ-ның, ал А.А. Баев – валиндік тРНҚ-ның алғашқы құрылымын анықтады.
1966 жылы – М. Ниренберг, С. Очоа және Х.-Г. Корана (M.Nirenberg, S. Ochoa, H.-G. Khorana) генетикалық кодты ашты.
1967 жылы – М. Геллерт (M. Gellert) ДНҚ фрагменттерін біріктіретін фермент – ДНҚ-лигазаны ашты.
1970 жылы – Г. Темин мен Д. Балтимор (H. Temin, D. Baltimor) онкогендік вирустардағы кері транскриптазаны (РНҚ-тәуелді ДНҚ-полимераза) ашты.
1972 жылы – П. Боэр, С. Коэн және П. Берг (P. Boyer, S. Cohen, P.Berg) ДНҚ клондарын жасау технологиясын жасады, ген инженериясының негізін құрады.
1972 жылы – Х.-Г. Корана (H.-G. Khorana) аланиндік тРНҚ генінің химиялық синтезін жүзеге асырды.
1975-1977 жылдары – Ф. Сангер (F. Sanger), сондай-ақ А.Максам мен У. Гилберт (A. Maxam, W. Gilbert) ДНҚ бастапқы құрылымын тез анықтау тәсілін ойлап тапты.
1976 жылы – Ф. Сангер (F. Sanger) φХ174 фагының ДНҚ нуклеидтік тізбегін анықтады.
1976 жылы – У. Гилберт (W. Gilbert) эукариот гендерінің мозаикалық құрылысын ашты.
1976 жылы – С. Ким, А. Рич және А. Клуг (S. Kim, A. Rich, A.Klug) тРНҚ-ның үшіншілік құрылымын анықтады.
5. ХХ ғасырдың аяғында молекулалық биологияның жетістіктері. Қазақстанда молекулалық биологияның дамуы
ХХ ғасырдың 70- ші жылдардың аяғы мен 80-ші жылдарында молекулярлық биология өзінің шарықтап даму сатысына өтеді. Осы уақытта сплайсинг механиздері ашылды,(В. Келлер және бас.), РНК- ферменттері (рибозимдер) ашылады және аутосплайсинг құбылысы түсіндіріледі (Т. Чек), генетикалық рекомбинация механизмдері және қозғалғыш генетикалық элементтер өте белсенді түрде зертеле басталады (Д.Хогнесс, Г. П. Георгиев), ферменттер мен биологиялық мембраналарды зерттеу жаңа деңгейге көтеріледі (Ю. А. Овчинников), жоғарғы организдердің геномдарының құрылысын анықтау жұмыстары басталады (оның ішінде адам геномыда кіреді), жаңа (гендік- инженерлік) биотехнологияның негізі жасалынады, жаңадан катализдік активті антитела (абзимдер) табылған және синтезделген, белоктар инженериясы саласы пайда болған.
ХХ-ғасырдың 80-жылдарындағы молекулалық биологияның шарықтап дамуы жаңа ғылым— биоинформатиканы (есептеу биологиясы, компьютерлық генетика), қалыптастырды. Ол молекулалық генетика мен информатиканың тоғысқан аймағынан пайда болды. Оның пайда болуына 1975— 1976 ж. Ф. Сангер мен А. Коулсон, сонымен қатар А. Максам және У. Гильберт жасаған ДНҚ-ның нуклеотидттік қатарын анықтаудың жылдам тәсілдері себеп болған. 1982 ж. нуклеотидтік қатарлардың банклері жасалған, онда әртүрлі организмдер ДНҚ-ның нуклеотидтік қатарлары туралы информация жинақталған.
ХХ-ғасырдың аяғындағы әртүрлі организдер ДНҚ-ның нуклеотидтік қатарларын анықтау (секвендеу) саласындағы жетістіктер (бірінші рет толық бактериялық геном 1995 ж. секвенделген, 1997 ж. — ашытқы геномы, 1998 ж. — нематод геномы, 2000 ж. — дрозофилла шыбыны геномы және толықтайға жақын адам геномы- секвенделген), ол геномиканы қалыптастырды. Геномика - бұл берілген организмнің гендер жиынтығын бір тұтас күйінде зерттейтін ғылым саласы. Онымен біруақытта кез-келген организмнің әртүрлі даму сатыларындағы функциялық қызмет атқаратын белоктардың толық жинағын зерттейтін протеомика деген ғылым саласы пайда болды.
XX ғасырдың 90 жылдары жоғары сатылы диплоидты ағзаларды клондау тәжірибелері сәтті аяқталып, 1997 ж. Үлыбританияда «Долли» атты қозы, 1998 ж. торай, ал 1999 ж. маймыл баласы дүниеге келді.
2001-2003 жылдары «Адам геномы» атты халықаралық ғылыми бағдарлама толық аяқталып, 2001 жылдан кейін адамзат постгеномдық дөуірге аяқ басты.
Молекулалық биологияның дамуына көптеген орыс және қазақ ғалымдары ат салысты, олардың арасынан А.А.Баев, А.Н.Белозерский, А.С.Спирин, В.А.Энгельгард, А.П.Георгиев, ТДарханбаев, МААйтхожин, Х.Жуматов т.б. есімдерді атауға болады.
Біздің отандас академик М.Ә.Айтхожин өзінің ұстазы А.А. Спиринмен бірлесіп 1964 жылы информасоманы-РНҚ және белоктардың ерекше формасынан тұратын жасушалық субмикроскопиялық бөлшектерді ашты. Осы ашылған жаңалықтар үшін ғалымдар Лениндік сыйлық (1976) иегерлері болды. Мұрат Әбенұлы Айтхожин дүниеден ерте озды (1987), алайда Қазақстанда біздің ұлы отандасымыздың атындағы Молекулалық биология және биохимия институтында зерттеулер жалғасуда.
1983 жылы құрылған Молекулалық биология және биохимия институты үлкен ғылыми орталыққа айналып, елімізде алғаш рет клеткалық және гендік инженерия, өсімдік геномы зерттеле бастады. Алматы мен Астанада биотехнологиялық орталықтар құрылды.
Ұсынылатын әдебиеттер тізімі:
Негізгі
1. Коничев А.С, Севастьянова Г.А. Молекулярная биология. – М.: ACADEMA, 2005.
2. Аношкина Е.В., Кондратенко Е.И., Ломтева Н.А. Молекулярная биология. – Астраханский университет, 2007.
3. Уотсон Дж. Молекулярная биология гена. М.: Мир, 1978.
4. Молекулярная биология: Структура и биосинтез нуклеиновых кислот (под ред. Спирина А.С.-М.: Высшая школа, 1990.
5. Молекулярная биология: Структура рибосом и биосинтез белка (под ред. Спирина А.С.). –М.: Высшая школа, 1996.
6. Степанов В.М. Молекулярная биология. Структура и функции белков.-М.:Высшая школа, 1996.
7. Альбертс Б., Брейм Д., Льюис Дж., Рефф М., Робертс К.,Уотсон Дж. Молекулярная биология клетки в 5-и томах.-М.:Мир, 1994.
8.Сингер М., Берг П. Гены и геномы. В 2-х томах.-М.: Мир,1998.
9.Уотсон Дж., Туз Дж., Куру Д. Рекомбинантные ДНК.-М.: Мир, 1986.
10.Льюин Б.Гены. –М.: Мир, 1987.11. Зенгбуш П. Молекулярная и клеточная биология в 3-х томах.-М.: Мир, 1982.
11. Эллиот В., Эллиот Д. Биохимия и молекулярная биология.-М.:НИИ Биомедицинской химии РАМН, 2000.
Қосымша:
- Овчиников Ю.А., Биорганическая химия.-М.: Просвещение, 1987.
- Malacinski G.M., Freifelder D. Essentials of Molecular Biology. London. Jones and Bartlett Publishers, 1998.
Тақырып: Нуклеин қышқылдарының генетикалық ролі. ДНҚ –ның қос спиральді моделінің үлгісі,. Молекулалық биологияның зерттеу тәсілдері
1.Нуклеин қышқылдарының генетикалық ролі
2. ДНҚ –ның қос спиральді моделінің үлгісі
3. Молекулалық биологияның зерттеу тәсілдері
1. Нуклеин қышқылдарының генетикалық ролі
ДНҚ-ның тұқымкуалаушылың қүбылысындагы рөлін айкындауда, Ф. Гриффитстің 1928 ж. пневмококк бактерияларында ашқан трансформация қүбылысының маңызы зор. Пневмококк бактериялары сүтқоректілер өкпесінің қабынуын (пневмонияны) қоздырып, олардың өліміне себепші болады. Сондықтан мұндай бактериялар патогенді немесе вирулентті болып саналады, себебі олардың полисахаридті кабығының шырышты бөлігі жұқтырылған жануар иммундық жүйесінің фагоциттеріне қарсы антизаттар (у) бөліп шығарады. Вирулентті бактериялар қоректік ортада тегіс шоғыр ( S-штамм) түзеді. Шырышты қабығы жоқ вирулентті емес бактериялар мутация арқылы пайда болады. Олар қоректік ортада кедір-бүдыр шоғыр (R-штамм) түзеді. Тышқандарға осындай бактерияларды енгізсе, онда олар фагоцитоз нәтижесінде бактериялық клеткаларды жойып, тірі қалады. Бірақ вирулентті S бактерияларымен инъекцияланған тышқандар өкпесінің қабынуынан өледі, өйткені бұл бактериялардың сыртын өздері синтездейтің шырышты қабық жабады. Ал алдын ала кыздыру арқылы өлтірілген S бактериямен (шырышты қабығынан айырылған) инъекцияланған тышқандар тірі қалады.
Ф. Гриффитс тышқандарға пневмококтың R –штамын және қыздыру арқылы капсуласынан айырылған S -штамын бірге инъекция лайды. Бүл арада күткен нәтиженің -тышқандардың тірі қалуының орнына, олардың барлығы өліп қалады (1-сурет).
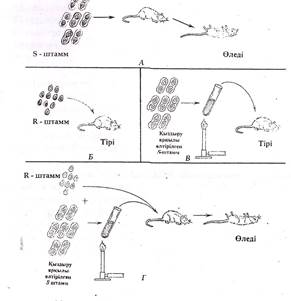
1-сурет.Трансформация құбылысы.
А-тышқанға вирулентті S-штамм енгізілді, тышқан өледі; Б-тышқанға авирулентті R-штамм енгізілді, тышқан тірі қалады; В-тышқанға қыздыру арқылы өлтірілген вирулентті S-штамм енгізілді, тышқан тірі қалады; Г-тышқанға авирулентті R және қыздыру арқылы өлтірілген вирулентті S-штаммдары бірге енгізілді, тышқан өледі.
Пневмониядан өлген тышқандардан шырышты қабығы бар S-вирулентті штамм бөлініп алынды. Демек, S -штамының вируленттік қасиетін анықтайтын зат -R штамға өтетіні анық болды.
Осыдан келіп, Гриффитс вирулентті емес R-штамм вирулентті S-штамға ауыса алады деген қорытынды жасады. Құбылыстыңөзі трансформация деп, ал бактерия ның қасиетін өзгертетін зат – трансформациялаушы фактор деп аталды.
Көп жылдар бойы трансформациялаушы фактор жөне оның субстанциясы жүмбақ болып келді. Тек 1944 ж. американ бактериологтері О. Эвери, К. Маклеод және М. Маккарти трансформациялаушы фактор, яғни тұқым қуалау қасиетін өзгерте алатын зат ДНҚ екенін атапкөрсетті. Олар өсіп жаткан - R бактериялар себіндісіне (культурасына) S-штамнан тазартылып алынған ДНҚ қосылса, кейбір R -бактериялар полисахаридті қабық түзетінін байқады. Кейін Эвери және оның қызметкерлері трансформациялаушы фактор дезоксирибонуклеаза ферментінің әсерінен жойылатынын накты деректерімен көрсетті, ал бүл ферменттің тек ДНК молекуласын ғана ажырататыны бұрыннан мәлім болатын. Бірақ олар ашкан жаңалықтың іргелі мән-мағынасы әр түрлі себептермен өз уақытында бағаланбады.
Біріншіден, ДНҚ-ның химиялың құрылымы айқын емес еді: ДНҚ - химиялық тұрғыдан жеткілікті түрде ұйымдастырылмаған қосылыс, сондықтан да ол ақпаратты өзіне сақтай алмайды, Екіншіден, ақуыздың құрылысы өте күрделі, сондықтан да сол кезде гендер ақуыздан түрады деген пікір қалыптасқан еді. Ақырында бактерия мен жоғары сатыдағы ағзалардың генетикалық ақпараттың жалпы принциптері бірдей деп қаралмады. Осыған байланысты бактерияларда тұқым қуалайтын зат ДНҚ, ал жануарлар мен өсімдіктерде басқа зат болар деген жорамал айтылды.
Тұкым қуалауда ДНК-ның басты рөл атқаратынын 1952 ж. А. Херши мен М. Чейз бұлтартпай дәлелдеп берді. Олар тәжірибені Т2 бактериофагына жүргізді. Бүл вирус ДНҚ-дан және ақуыз қабығынан тұрады. Фагтың ақуызды қабығы радиоактивті күкіртпен (S35), ал ДНҚ- сы радиоактивті фосформен (Р32) белгіленді. Бактерияны радиоактивті элементтермен, белгіленген фагтармен жұқтырғанда фосфордың клеткаға енгені, ал күкірттің оның сыртында қалғаны байқалды. Бактерия клеткаларында көптеген жаңа пісіп-жетілген фагтар пайда болды. Бұдан бактерияға фаг ДНҚ-сы өтеді, жаңадан түзілген фагтардың барлық қасиеттері ДНҚ-ның. бақылауында болады деген қорытынды жасауға болады.
Қазіргі кезде эукариоттарда бактериялардың трансформациясы айнымай байқалады, бірақ ол тарихи сипаттағы себептерге байланысты трансфекция деп аталады. Эукариоттарда тимидинкиназа ферментінің синтезделуін TК-гені басқарады. ТК-гe ні жоқ эукариот клеткалары тимин жоқ болса өспейді. Осындай клеткаларға ТК-гені орналасқан ДНҚ молекуласын қосқанда олардың кейбіреулерінің өсіп, жеке шоғыр қүрағаны байқалды.
Кейбір вирустар ДНҚ-дан емес, рибонуклеин қыш қылынан (РНҚ-дан) қүралған, Химиялық күрылысы жағынан РНҚ-ның ДНҚ-дан аздаған айырмашылығы болғанымен, мүндай вирустарда ол генетикальқ материал ретінде пайдаланылады. Сонымен, прокариоттардың басым көпшілігіне және барлық эукариоттарда генетикалық ақпараттың іске асуын ДНҚ, ал кейбір вирустарда РНҚ бақылайды деп қорытынды жасауға болады.
2. ДНҚ –ның қос спиральді моделінің үлгісі.
1953 ж. Дж.Уотсон және Ф. Крик ДНК құрылымының моделін ұсынған. Моделді жасағанда ғалымдар оларға белгілі төрт ғылыми мағұлыматтарға сүйенді:

