




Для обсуждения роли количества и качества питательных веществ в жизнедеятельности животных рассмотрим их пищевые режимы и пищевые специализации. Под пищевым режимом понимается своеобразие, природа
85
пищевого материала, являющегося источником энергии для того или иного вида животных. Различают три основных типа пищевых режимов: зоофагия, фитофагия и детритофагия. Первый основан на использовании в пищу животных организмов и продуктов их жизнедеятельности. В зависимости от вида животной пищи выделяют следующие группы зоофагов: питающиеся живыми тканями (биофаги), трупами (некрофаги), экскрементами (копрофаги).
В качестве особых случаев зоофагии можно рассматривать питание такими специфическими материалами животного происхождения, как воск, шелк, шерсть. Подобные варианты питания интересны для изучения природы усваиваемых продуктов и механизма их расщепления в организме.
Фитофагия основана на использовании для питания растительных организмов и их продуктов. Число видов-фитофагов огромно. У цветковых (покрытосеменных) растений в пищу употребляется все — листья, лепестки, нектар, корни. Широко распространено питание древесиной, или ксилофагия.
Разлагающиеся растения, или детрит, и продукты их разложения служат пищей для видов-детритофагов, среди которых особенно много почвенных форм. Принимая активное участие в разложении лесной подстилки, детритофаги играют большую роль в обмене веществ между почвой и растительностью. Вместе с детритофагами действуют животные, питающиеся микроорганизмами; отделить одну группу от другой довольно трудно. Эти животные имеют небольшие размеры и способны поедать бактерии и грибы, но не сам детрит.
Детритофаги населяют наземные местообитания всех типов. Как правило, они представлены очень большим числом видов и достигают высокой численности. Например, в лесах умеренной зоны на 1 м2 почвы можно обнаружить около 1000 видов животных, при этом численность нематод и простейших может превышать 10 млн, ногохвосток и почвенных клещей — 100 тыс., других беспозвоночных —около 50 тыс. Удельный вес
86
микро-, мезо- и макрофауны в наземных сообществах изменяется в меридиональном направлении —от тропиков к полюсу (рис. 5.3). Микрофауна играет существенную роль в богатых органикой почвах северных лесов, тундры и полярной пустыни.
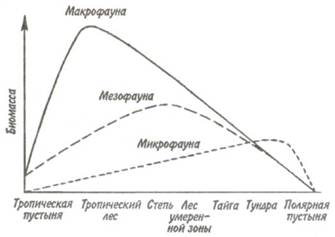
Рис. 5.3. Типы широтных изменений вклада макро-,
мезо- и микрофауны в процессе
разложения биомассы в наземных экосистемах
(по: Бигон, Харпер, Таунсенд, 1989).
Большое количество органического материала в почвах стабилизирует режим увлажненности последних и обеспечивает благоприятные микроусловия для существования простейших, нематод и коловраток, которые обитают в интерстициальной водной пленке. Этих животных очень мало в прогретых, сухих минерализованных почвах тропиков. Обогащенные органическим веществом почвы в лесах умеренной зоны занимают промежуточное положение. В них поддерживается высокая численность популяций почвенных клещей, ногохвосток и энхитреид, относящихся к мезофауне. Численность большинства других групп животных по направлению к более засушливым тропикам уменьшается; там их замещают термиты.
От пищевых режимов следует отличать пищевую специализацию, т. е. круг пищевых материалов, используемых данным видом. Он может быть широким или строго ограниченным. В зависимости от степени ограниченно-
87
сти пищевого рациона виды-гетеротрофы распадаются на три экологические группы: полифаги, олигофаги и монофаги.
Обширную группу животных и растений используют в качестве пищи полифаги. К ним относятся многие виды животных: тараканы, сверчки, бурый медведь и др. Большинство паразитов специализировано в отношении выбора хозяина, но некоторые из них могут быть полифагами. Для олигофагов спектр пищевых объектов ограничен сравнительно узкими рамками. Паразиты, хищники и фитофаги часто питаются представителями только определенных родов или семейств из мира животных или растений. У монофагов набор объектов питания ограничен одним видом или несколькими близкими видами одного рода растений или животных. Среди растительноядных насекомых монофагами являются некоторые виды, используемые для уничтожения сорняков.
Червец, завезенный из Индии в Австралию для борьбы с сорными кактусами-опунциями, оказался настолько специалиализированным монофагом, что сумел справиться лишь с одним видом опунции. Другой пример также связан с Австралией. На Австралийском континенте в ппериод бурного развития скотоводства и освоения обширных территорий под пастбища местная фауна жуков-навозников, способных питаться только экскрементами сумчатых, оказалась неприспособленной к питанию навозом овец и крупного рогатого скота. Навоз накапливался в огромных количествах на поверхности земли, затрудняя рост травы и снижая продуктивность пастбищ. Для ликвидации последствий нарушенного экологического равновесия пришлось интродуцировать в Австралию несколько видов навозников из Южной Африки. Интродуцированные навозники, усваивающие экскременты копытных, способствовали освобождению пастбищ от навоза и удобрению почвы продуктами его переработки.
Границы между моно-, олиго- и полифагами, разумеется, условны. Есть немало животных, которых трудно отнести к той или иной группе, однако сам факт существования у всех животных пищевого предпочтения и специализации по отношению к определенному источнику пищи не подлежит сомнению. Пищевая специали-
88
зация обусловливается различными причинами: способом добывания пищи, быстротой реакций и др.
У хищников набор пищевых объектов определяется в первую очередь способом ловли добычи и местом охоты. Так, хищные гусеницы не обладают быстрыми двигательными реакциями, поэтому их добычей могут быть только неподвижные или малоподвижные насекомые. Однако нападение богомола провоцируется движением объекта в его поле зрения, в связи с чем богомолы не могут питаться неподвижными животными — червецами или щитовками. Различный способ охоты свойствен также разным кошачьим. Так, гепард догоняет свою жертву, а леопард подстерегает ее, затаившись на дереве.
Добывание пищи для животного имеет определенную «стоимость», измеряемую затратами времени и энергии. Ясно, что животное может существовать только в том случае, если получаемая с пищей энергия превышает затраты на питание и у него остается время на другие формы активности, в том числе на размножение. В процессе естественного отбора у животных формируется оптимальная стратегия добывания пищи, позволяющая увеличить разность между поступающей с пищей энергией и необходимыми ее затратами на питание. Оптимизируются такие параметры, как скорость и характер передвижения, способы поиска корма, тип и размеры добычи, пищевой рацион, размеры кормового участка и др. Теория оптимального добывания пищи имеет важное значение в экологии животных.
У животных известно два основных типа питания. Пассивное питание характерно в основном для сидячих водных животных, которые питаются, отфильтровывая из воды взвешенные пищевые частицы (губки, двустворчатые моллюски и др.). К этому же типу питания относится всасывание питательных веществ поверхностью тела (многие простейшие и некоторые паразиты). Активное питание свойственно большинству животных и требует их передвижения во внешней среде для добывания корма. Пассивное питание неизбирательно, а активное, как правило, отличается избирательностью, хотя существуют
89
активно питающиеся животные с неизбирательным питанием (грунтоеды, усатые киты, отдельные виды рыб и др.).
Активно питающихся животных разделяют на две большие группы — «жнецы» (собиратели) и «охотники». Первые используют пищу, имеющуюся в изобилии, а вторые специализированы на добывании более редкого корма. Жнецы — это не только растительноядные животные; к ним относятся и многие «мирные» зоофаги (планктоноядные рыбы, усатые киты, многие насекомоядные птицы и звери и др.), использующие легко добываемую пищу, которую не нужно долго искать. Группа охотников включает не только хищников. Сюда входят те фитофаги, которые вынуждены отыскивать пищу (например, питающиеся семенами и плодами).
Жнецы и охотники не полностью изолированные группы, между ними существуют постепенные переходы. Однако такое разделение на группы позволяет отчетливо выявить многие экологические закономерности. Так, для охотников характерно сложное территориальное поведение, а жнецы редко защищают свою территорию, но среди них чаще встречаются виды, ведущие сложный общественный образ жизни.
Для жнецов характерны две формы питания. При пастьбе животные уничтожают только часть имеющихся в данном месте кормов и постепенно перемещаются на новые участки. Выедание отличается тем, что корм уничтожается весь или почти весь, и только после этого животные мигрируют в другие места.
Охотники по формам питания разделяются на три типа. Многие хищники («засадники») подкарауливают свою добычу и нападают на нее неожиданно, обычно из укрытия (богомолы, щуки, ястребы, кошки и пр.). Другие животные («искатели»), например насекомоядные птицы, проводят значительную часть времени в поисках добычи, но преследуют, схватывают и поедают ее довольно быстро, расходуя на это сравнительно мало энергии. Охотники третьего типа («преследователи») затрачивают немного времени на поиски добычи, но овладение ею требует от них значительных усилий (многие крупные хищники, соколы и т. п.). Конечно, некоторые животные могут использовать все три способа охоты в зависимости от ситуации, возможны также промежуточные варианты.
Способ добычи ограничивает круг возможных источников пищи для хищника, однако эти факторы не приводят к строгой монофагии. Абсолютная монофагия свойственна некоторым насекомым-парализаторам. В этом
90
случае, в связи со сложностью инстинктивных операций, направленных на ^обездвиживание жертвы, необходима очень тонкая и точная подгонка поведения хищника к строению тела жертвы.
Добычей лангедокского сфекса служат неповоротливые крупные бескрылые кузнечики эфиппигеры. Сфекс, схватив свою жертву за грудной щит, наносит ей два последовательных укола жалом, сначала снизу в грудь, чтобы поразить средний и задний нервные узлы, а затем в горло, чтобы поразить переднегрудной узел. Песчаная аммофила при парализации гусеницы последовательно жалит ее снизу в каждый сегмент, постепенно перехватывая ее челюстями и передвигаясь от головы к заднему концу.
Ярко выраженная пищевая специализация обнаруживается у многих паразитических животных. Факторами, обеспечивающими и поддерживающими узкую олигофагию, могут быть особенности поведения паразита и его способность развиваться в строго определенных хозяевах. Адаптированность каждого вида паразита к своему хозяину проявляется, во-первых, в морфологических изменениях, характерных для паразитического образа жизни, во-вторых, в развитии физиологических реакций подавления иммунитета и, в-третьих, в синхронизации циклов развития паразита и хозяина.
Пищевая специализация фитофагов обычно основана на их способности реагировать на определенные химические компоненты растений. Стимулами, привлекающими или отталкивающими животных, могут быть различные органические соединения, которые содержатся в растительном субстрате, но которые, как правило, не имеют питательной ценности. Чаще всего функцию этих стимулов выполняют гликозиды или терпены. Химические вещества, привлекающие животных, называются аттрак тантами, а отталкивающие— репеллентами. Растение употребляется в пищу, если оно содержит необходимые пищевые аттрактанты и не имеет достаточно сильных репеллентов. Обогащенная аттрактантами пища сильнее привлекает фитофагов и поглощается ими в большем
91
количестве. Поэтому существует прямая связь между степенью привлекательности пищи и ее «оптимальностью», т. е. способностью поддерживать высокие темпы роста животных при минимальной смертности. «Неоптимальная» пища может обладать столь же высокой питательной ценностью, но она лишена аттрактантов или имеет сильные репелленты и поэтому поглощается в меньшем количестве. Понятно, что при питании «неоптимальной» пищей рост и развитие животных замедляется, а смертность возрастает.
Когда пищевые аттрактанты широко распространены в мире растений и нет достаточно сильных репеллентов, мы имеем дело с полифагией. Пищевая специализация животных-олигофагов может быть связана с ограниченным распространением аттрактантов или, наоборот, с широким распространением и аттрактантов и репеллентов. Наконец, строгая монофагия обусловливается аттрактантами, которые свойственны только одной небольшой группе растений.
Когда пищевые аттрактанты свойственны лишь ограниченному кругу растений, то возникает олигофагия. Так, у гусениц и бабочек-белянок специализация на питании крестоцветными несомненно связана с привлекающим действием гликозидов горчичного масла.
Помимо узкой олигофагии выделяют также аномальную олигофагию, т. е. питание растениями нескольких видов неблизкородственных родов. Аномальным олигофагом является бабочка тутовый шелкопряд, гусеницы которой питаются белой шелковицей и маклюрой. Для гусениц этого вида известен ряд привлекающих веществ, которые по своему действию делятся на три группы: дистантные пищевые аттрактанты, контактные пищевые аттрактанты, вещества, вызывающие заглатывание пищи. Первая группа веществ привлекает гусениц на расстоянии и вызывает их передвижение по направлению к источнику корма. Контактные пищевые аттрактанты вступают в действие при непосредственном соприкосновении гусениц с кормом и способствуют отгрызанию кусочка листа. Для проглатывания его необходимо присутствие третьей группы веществ. Таким образом, аномальная олигофагия возникает в том случае, когда питание
92
стимулируется большой группой различных веществ', которые редко присутствуют вместе в одном и том же растении.
Различия между этими тремя экологическими группами фитофагов носят не качественный, а скорее количественный характер (рис. 5.4). Монофаги и в меньшей степени олигофаги связаны с определенной и ограниченной группой кормовых растений. Потребление этих растений обеспечивает оптимальную скорость развития, плодовитость и прочие процессы жизнедеятельности. Поэтому в данном случае лишь количество пищи служит лимитирующим фактором. Иначе обстоит дело с полифагами. Здесь ограничивающим фактором становится качество пищи, т. е. содержание в ней определенных веществ, необходимых для осуществления различных процессов жизнедеятельности.
В зависимости от стадии жизненного цикла полифаг оказывает предпочтение тем или иным ее видам. Так, самкам шотландской куропатки перед размножением

Рис. 5.4. Эффективность поедания кормовых растений
тремя группами насекомых-фитофагов.
Кормовые растения: 1 - отвергаемые, 2 - непривычные,
3 - замещающие, 4 - оптимальные.
93
необходимы такие элементы, как кальций, фосфор и азот. Поэтому птицы выбирают в качестве корма вереск, богатый этими элементами, не трогая растений, которые содержат их в недостаточном количестве.
Чем уже спектр потребляемых организмом пищевых ресурсов, тем теснее должна быть пространственная связь этого организма со скоплениями его ресурсов или же тем больше должны быть затраты времени и энергии на поиски необходимых ресурсов.
Наличие и доступность многих пищевых ресурсов зависят от времени года. Хорошим примером могут служить заросли дикой малины в лесах умеренного пояса. Зимой растения малины представляют собой скопление мелких сучьев, но весной на них во множестве появляются молодые, богатые белком почки, а затем листья. Зацветание влечет за собой кратковременное выделение нектара, а нектар — это совершенно новый ресурс, существующий, правда, лишь во время цветения. Со временем завязываются плоды, и плоды эти, созревающие и созревшие, представляют собой новый обильный источник корма. Все эти сезонные образования могут являться надежными ресурсами либо для растительных полифагов, которые в состоянии переключиться на другую пищу, когда «сезон малины» минует, либо для монофагов, у которых весь период активной жизни втиснут в соответствующий сезон и которые проводят всю остальную часть года в спячке, или диапаузе, не нуждаясь в пище.
94
Т е м а 6
БИОТИЧЕСКИЕ ФАКТОРЫ
Под биотическими факторами понимают различные формы взаимодействия между особями и популяциями. Биотические факторы подразделяются на две группы: межвидовые и внутривидовые взаимодействия.
ГОМОТИПИЧЕСКИЕ РЕАКЦИИ
Под гомотипическими реакциями понимают взаимодействия между особями одного вида. Жизнедеятельность животных и растений при совместном обитании в значительной степени зависит от численности и плотности популяции. В связи с этим большое экологическое значение имеют явления, связанные с так называемыми эффектами группы и массы.
Эффект группы — это влияние группы как таковой и числа индивидуумов в группе на поведение, физиологию, развитие и размножение особей, вызванное восприятием присутствия особей своего вида через органы чувств.
Многие насекомые (сверчки, тараканы, саранчовые и др.) в группе имеют более интенсивный, чем при жизни поодиночке, метаболизм, быстрее растут и созревают. Баклан —главный производитель гуано в Перу, может существовать лишь при условии, если в его колониях насчитывается не менее 10 000 особей и на 1 м2 приходится 3 гнезда. При совместной жизни легче искать и добывать пищу, а также защищаться от врагов. Объединенные в стаю волки способны убивать добычу более крупных размеров, чем действуя в одиночку. Бизоны, мускусные быки и другие жвачные успешнее обороняются от хищников, если они объединены в стада.
Нередко эффект группы проявляется уже при совместном существовании двух животных. Он может приводить не только к положительным, но и к отрицательным
95
результатам. Например, рост головастиков в группе замедляется. Как правило, при небольшой численности группы преобладают положительные эффекты, а при избыточной плотности животных доминируют отрицательные явления. Так, в перенаселенных группах домовых мышей падает плодовитость и даже совсем прекращается размножение. У мучного хрущака при высокой плотности популяции намного увеличивается доля бесплодных яиц.
Групповой эффект наиболее ярко выражается при наличии у животных явления фазности, т. е. способности вида существовать одновременно в двух формах — в форме одиночных особей и в форме групп. Эффект группы наглядно виден у мигрирующих саранчовых, которые при изолированном воспитании представляют собой одиночную фазу (солитария), окрашенную в зеленый цвет, а при скученном воспитании возникает стадная фаза (грегария), отличающаяся контрастными черными пятнами на оранжевом или светло-коричневом основном фоне.
Эффект массы, в отличие от эффекта группы не связан с восприятием особями одного вида присутствия друг друга. Он выливается изменениями в среде обитания, происходящими при увеличении численности особей и плотности популяции. Как правило, эффект массы отрицательно сказывается на плодовитости, скорости роста, длительности жизни животных. Например, при развитии популяции мучного хрущака в муке постоянно накапливаются экскременты, линочные шкурки, что приводит к ухудшению муки как среды обитания. Это вызывает падение плодовитости и повышение смертности в популяции жуков.
В природе эффекты группы и массы далеко не всегда легко различить, поскольку они нередко проявляются одновременно. Исключительно важную роль играют групповые и массовые эффекты в динамике численности популяций, выступая в ряду так называемых зависящих от плотности факторов, которые регулируют численность популяции по принципу обратной связи.
96
Здесь следует упомянуть принцип Олли: для каждого вида существует оптимальный размер группы и оптимальная плотность популяции. Как перенаселенность, так и недоселенность (или отсутствие агрегации) могут оказывать неблагоприятное влияние. Это положение подтверждено многочисленными наблюдениями и экспериментами. Известно, например, что для успешного выживания стадо слонов должно состоять по крайней мере из 25 особей, а стадо северного оленя — минимум из 300 голов. Гнездящиеся совместно чайки и другие птицы часто не могут успешно размножаться, если колония слишком мала. Однако и очень большая численность группы для них оказывается неблагоприятной. Таким образом, действие принципа Олли объясняется совместным влиянием эффектов группы и массы.
К гомотипическим реакциям кроме группового и массового эффектов относится еще одна форма взаимодействия между особями одного вида — внутривидовая конкуренция. Сам термин появился сравнительно недавно. Раньше в том же значении использовался термин «борьба за существование», предложенный Ч. Дарвином, т. е. борьба за возможность выжить, для чего необходима энергия, получаемая растениями в виде солнечного света, а животными в виде различных пищевых материалов. Поэтому в борьбе за овладение источниками энергии происходит напряженная конкуренция, возникает соперничество между особями одного вида. В отношении всех видов конкуренции существует правило: чем более совпадают потребности конкурентов, тем острее конкуренция. Следовательно, внутривидовая конкуренция острее межвидовой, конкуренция между особями одной популяции острее, чем между особями разных популяций, и т.д. Победивший в конкурентной борьбе имеет возможность оставить потомство, а, следовательно, передать свои гены по наследству. Уступивший в конкурентной борьбе погибает или, точнее, элиминируется. Таким образом, основной результат, к которому приводит внутривидовая конкуренция,— дивергенция особей, завершающаяся формированием популяций (рис. 6.1).
97

Перечеркнуты кружки, соответствующие особям, которые не оставили потомства.
Различают две основные формы конкуренции — прямую и косвенную. Прямая конкуренция, или интерфе ренция, осуществляется путем прямого влияния особей друг на друга, например, при агрессивных столкновениях между животными или при выделении токсинов (аллелопатия) у растений и микроорганизмов. Интерференция у животных не обязательно заключается во взаимной агрессивности. Она может возникать, например, когда одни животные своим поведением лишают других доступа к пище, укрытиям, к местам для размножения и т. д.
Косвенная конкуренция не предполагает непосредственного взаимодействия между особями. Она происходит опосредованно — через потребление разными животными одного и того же ресурса, который обязательно должен быть ограниченным. Поэтому такую конкуренцию обычно называют эксплуатационной. В качестве причи-
98

 ны, вызывающей конкуренцию, может выступать любой лимитированный ресурс, а также места для размножения, укрытия и т. д. При эксплуатационной конкуренции преимущество получают животные, способные быстрее и эффективнее использовать ресурсы, являющиеся предметом конкуренции, и уменьшать их количество, достающееся другим животным.
ны, вызывающей конкуренцию, может выступать любой лимитированный ресурс, а также места для размножения, укрытия и т. д. При эксплуатационной конкуренции преимущество получают животные, способные быстрее и эффективнее использовать ресурсы, являющиеся предметом конкуренции, и уменьшать их количество, достающееся другим животным.
Внутривидовая конкуренция нередко проявляется в территориальности—охране отдельными животными или их группами индивидуальных участков. Обычно охраняется кормовая территория. Это характерно для некоторых видов насекомых, рыб, многих видов ящериц, млекопитающих и подавляющего большинства видов птиц. Существуют также гнездовые и брачные территории, появление которых обусловлено наличием конкурентной борьбы за места размножения или за брачного партнера. Конкуренция за некоторый ограниченный ресурс обычно приводит к более равномерному пространственному размещению животных. У животных с общественным поведением конкуренция нередко выражается в возникновении иерархии доминирования, позволяющей упорядочить распределение ресурсов между животными в группе.
Поскольку основным объектом внутривидовой конкуренции служит энергия, то среди растений наиболее острая борьба идет за свет. При слишком большой густоте произрастания растения затеняют друг друга. Конкуренция выражается также в гибели определенного количества растений, в результате чего плотность последних снижается. Это явление хорошо выражено у некоторых культур (морковь, свекла). Оно наблюдается и в лесу, где молодых деревьев гораздо больше, чем старых.
Конкуренция за свет влияет на форму деревьев: у них совершенно различный вид в зависимости от того, растут ли они в лесу или поодиночке. Например, у дуба, если он растет отдельно, крона шарообразной формы, так как нижние боковые ветки получают достаточное количество света, чтобы развиваться не хуже верхних. В лесу же, напротив, листва нижних ветвей затенена вышерасположенными ветвями как этого дерева, так
99
и соседних растений. Происходит естественный процесс отмирания и опадения ветвей, листья которых получают мало света и у которых баланс ассимиляции и дыхания отрицательный. По мере роста в высоту опадение нижних ветвей происходит быстрее, и дерево принимает лесную форму, отличающуюся прямым цилиндрическим стволом, который на большом протяжении лишен ветвей.
Конкуренция протекает, но не по принципу «включена-выключена». Напротив, ее уровень непрерывно меняется в зависимости от конкретных условий. Так, в экологическом вакууме конкуренция слабо выражена или отсутствует, а в насыщенной среде она играет большую роль во взаимоотношениях между особями.
ГЕТЕРОТИПИЧЕСКИЕ РЕАКЦИИ
Гетеротипические реакции — это взаимодействия между особями, принадлежащими к разным видам. Две популяции либо влияют, либо не влияют друг на друга. Если влияние есть, то оно может быть благоприятным или неблагоприятным. Для рассмотрения гетеротипических реакции спекшим таблицу, где символами обозначены возможные взаимодействия.

