




Выше было приведено несколько примеров, показывающих действие температуры окружающей среды на животных. При этом нас интересовало, какое влияние оказывает изменение температуры на рост и развитие особей. Очевидно, что действие температуры на уровне популяции или вида скажется на значениях иных параметров или свойствах животных или растений. Рассмотрим влияние, которое оказывают абиотические факторы на жизнедеятельность животных и растений на уровне особи, популяции, вида и экосистемы.
УРОВЕНЬ ОСОБЕЙ
На уровне особи абиотические факторы оказывают влияние прежде всего на поведение животных, изменяют трофические отношения со средой и характер метаболизма, воздействуют на способность к размножению и плодовитость, в значительной степени определяют развитие, скорость роста и продолжительность жизни животных и растений. Существует два основных способа адаптации особи к изменениям абиотических факторов: поведенческие реакции и физиологическая перестройка организма.
Очевидно, что физиологические механизмы должны соответствовать тем экологическим условиям, в которых существуют организмы. Их физиология и экология оказываются в процессе эволюции тесно связанными. Именно в результате этого воздействие абиотических факторов на различные стороны жизнедеятельности организмов носит приспособительный характер, т. е. вызывает у них адекватную реакцию. Иначе говоря, в процессе эволюции образуются адаптации по отношению к экологическим факторам.
У растений существуют различные механизмы адаптации. Они затрагивают строение, развитие и обмен ве-
67
ществ. Модифицирующее действие освещенности местообитания на рост и морфогенез может быть проиллюстрировано динамикой роста двухлетних дубов Quercus robur в зависимости от относительной освещенности летом (рис. 4.1). Так называемые «световые» и «теневые» листья отличаются по целому ряду показателей: площади листовой пластинки, количеству клеток, густоте жилкования и т. д. На расположение и форму листьев оказывает влияние множество факторов, в том числе освещенность, наличие воды, доминирующие ветры и растительноядные животные. При росте в тени многие растения дают более крупные и менее рассеченные листья, чем при росте на ярком свету. Теневыносливые растения подлеска обычно характеризуются более крупными и более простыми по форме листьями, чем виды, образующие полог леса.
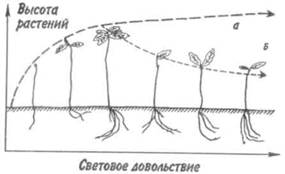
Рис. 4.1. Зависимость роста двухлетних дубов от
относительной освещенности летом (из Лархера, 1978).
При низком световом довольствии преобладает
стимулирующее действие света на рост (а), при большем
освещении — тормозящее действие (б).
Адаптации, возникающие к самым разнообразным факторам среды, распространены очень широко. Каждый конкретный организм обладает уникальным коадаптированным комплексом физиологических, поведенческих и экологических особенностей, дополняющих друг друга и способствующих более успешному его размножению.
68
 Подобное «созвездие» адаптации называют адаптивным комплексом, или оптимальной композицией. Для иллюстрации совершенства и слаженности адаптивного комплекса воспользуемся примером Э. Пианка.
Подобное «созвездие» адаптации называют адаптивным комплексом, или оптимальной композицией. Для иллюстрации совершенства и слаженности адаптивного комплекса воспользуемся примером Э. Пианка.
Рогатая ящерица Phrynosoma platyrhinos, обычное животное пустыни Нового Света, специализируется на питании муравьями и обычно кроме них ничего не ест. Поскольку муравьи—животные мелкие и содержат много неусвояемого хитина, ящерица должна поедать их в большом количестве. Особям любого вида, специализирующегося на питании муравьями, необходим большой, относительно размеров тела, желудок. Масса желудка рогатой ящерицы, выраженная в долях массы тела, составляет значительную величину (13%) по сравнению с массой желудка других видов ящериц. Обладание таким объемистым кишечником приводит к изменению формы тела (тело этих животных напоминает бочонок), что в свою очередь снижает скорость их движения и ограничивает способность удирать от хищников. Поэтому естественный отбор способствовал формированию на поверхности тела рогатой ящерицы шипов и переходу к скрытному образу жизни, хотя другим видам ящериц свойственны гладкая поверхность и высокая подвижность, что позволяет им быстро скрываться от врагов.
Вероятность быть съеденным хищником возрастает тогда, когда животное в поисках пищи пребывает длительное время на открытых пространствах. При этом снижение подвижности может оказаться и выгодным, так как движение привлекает внимание хищника и уменьшает преимущества, даваемые покровительственной формой и криптической окраской тела. Сниженная подвижность, несомненно, отчасти объясняет большую изменчивость температуры тела у рогатой ящерицы, которая значительно превышает таковую у всех других видов ящериц.
Рогатая ящерица активна в течение более продолжительного времени по сравнению с другими видами. Значительные колебания температуры ее тела вызваны как большой продолжительностью периода активности, так
69
и редкими перемещениями животных из тени и обратно. Очевидно, у рогатой ящерицы имеется больше времени для такого вида активности, как питание, а видам, специализирующимся на питании муравьями, всегда требуется для этого много времени. Специализация на питании муравьями экономически оправдана тем, что последние образуют четко выраженные скопления в пространстве и, следовательно, являются концентрированным источником пищи. Для того чтобы извлечь пользу из этой сконцентрированной, но не очень питательной пищи, у рогатой ящерицы в процессе эволюции выработался уникальный комплекс адаптации: это и объемистый желудок, и шипы на поверхности тела, продолжительный период активности и ослабленная терморегуляция (эвритермность).
Столь велико совершенство связанных друг с другом поведенческих реакций и физиологических перестроек, обеспечивших эволюционный успех рогатой ящерицы и именуемых адаптивным комплексом. Необходимо подчеркнуть, что адаптивный комплекс — явление видовое, так как все особи данного вида в равной степени обладают присущим ему набором поведенческих и физиологических реакций. Изучение одной, отдельно взятой особи позволяет обнаружить наличие адаптивного комплекса. В то же время другие эффекты, вызываемые действием абиотических факторов, проявляются только на популяционном уровне.
ПОПУЛЯЦИОННЫЙ УРОВЕНЬ
На уровне популяции абиотические факторы воздействуют на такие параметры, как рождаемость, смертность, средняя продолжительность жизни особи, скорость роста популяции и ее размеры, нередко являясь важнейшими причинами, определяющими характер динамики численности популяции и пространственное распределение особей в ней. Популяция может приспосабливаться к изменениям абиотических факторов, во-первых, меняя характер своего пространственного
70
 распределения и, во-вторых, путем адаптивной эволюции.
распределения и, во-вторых, путем адаптивной эволюции.
Избирательное отношение животных и растений к факторам среды порождает избирательность и к местообитаниям, т. е, экологическую специализацию по отношению к участкам ареала вида, которые он пытается занять и заселить. Участок ареала, занятый популяцией вида и характеризующийся определенными экологическими условиями, называется стацией. Выбор стации, как правило, определяется каким-нибудь одним фактором; это может быть кислотность, соленость, влажность и т. д.
Для эвритермных видов характерна зональная смена стаций, т. е. строго направленное изменение стаций при переходе вида из одной природной зоны в другую: при передвижении к северу избираются более сухие, хорошо прогреваемые открытые стации с разреженным растительным покровом, часто располагающиеся на легких песчаных или каменистых почвах; при передвижении к югу тот же вид заселяет более увлажненные и тенистые стации с густым растительным покровом и с глинистыми почвами (рис. 4.2). На приведенной диаграмме по характеру растительного покрова и микроклимату все стации разбиты на три экологические группы — ксеро-

Рис. 4.2. Диаграмма зональной смены стаций
(по Бей-Биенко, 1966).
Объяснения в тексте.
71
фитные, мезофитные и гигрофитные. Смещение популяций вида в более влажные стации при продвижении к югу показано косыми стрелками. При этом влаголюбивые популяции лесной и отчасти лесостепной зон лишаются возможности проникнуть в южные районы, так как более влажные, чем гигрофитные, стации физически и экологически немыслимы.
Вертикальная смена стаций аналогична зональной, но проявляется в горных условиях. Наиболее обычной ее формой служит переход популяций на более ксерофитные стации по мере повышения уровня их местообитания. Например, серый кузнечик в лесном поясе Кавказа держится на мезо- и гигрофитных стациях, а в альпийском поясе — на ксеро- и мезофитных.
Как можно видеть из закономерности смены стаций, важным экологическим фактором, определяющим выбор местообитаний наземных животных и растений, является влажность воздуха.
Стациальная приуроченность мокриц связана с содержанием в воздухе водяного пара Они многочисленны по берегам морей, где воздух насыщен влагой, и там живут открыто. В высокогорных районах с сухим воздухом мокрицы большую часть времени проводят под камнями и корой деревьев.
Мокрица Lygia oceanica обитает по берегам морей. Дневное время мокрицы проводят в укрытии. Но когда температура воздуха повышается до 20 °С снаружи и до 30 °С под галькой, они покидают свои укрытия и выползают на скалы, обращенные к солнцу. Причина такого перемещения состоит в том, что данный вид, очень плохо приспособленный к наземному местообитанию, имеет легкопроницаемую кутикулу. Когда влажность воздуха низкая, мокрица теряет много воды путем испарения, что и происходит на скалах, находящихся под солнцем. Интенсивное испарение снижает температуру тела животного, которая при нахождении его на скале равна 26 °С (рис. 4.3). Если же мокрица продолжает прятаться под галькой, где относительная влажность близка к 100%, а испарение равно нулю, температура тела достигает 30 °С.
Распределение стаций в водной среде обусловлено другими факторами, в частности кислотностью. Кислые
72

Рис. 4.З. Смена стаций мокрицей Lygia oceanica
в зависимости от температуры воздуха (из Дажо, 1975).
Объяснения в тексте.
воды торфяников способствуют развитию сфагновых мхов, но в них совершенно нет двустворчатых моллюсков-перловиц. Чрезвычайно редки в них и другие виды двустворчатых моллюсков, что связано с отсутствием в воде извести. Рыбы выносят кислотность воды в пределах рН от 5 до 9. При рН ниже 5 можно наблюдать их массовую гибель, хотя отдельные виды приспосабливаются и к среде, значение рН которой доходит до 3,7. Продуктивность пресных вод, имеющих кислотность менее 5, резко понижена, что влечет за собой значительное сокращение уловов рыбы.
Другой важный фактор, лимитирующий распределение водных животных и растений,— это соленость воды. Многие крупные таксономические группы в ранге типа (иглокожие, кишечнополостные, мшанки, губки, кольчатые черви и др.) все или почти все являются морскими.
73
Часто лишь незначительные сдвиги в концентрации солей в воде сказываются на распределении близкородственных видов (рис. 4.4). Численность обитателей солоноватых вод очень велика, но их видовой состав беден, поскольку здесь могут жить только эвригалинные виды как пресноводного, так и морского происхождения. Например, озеро с соленостью, изменяющейся от 2 до 7°/00 , населяют пресноводные рыбы, такие как карп, линь, щука, судак, хорошо переносящие небольшую соленость, и морские, например кефаль, которые терпимо относятся к недостаточной солености.

Рис. 4.4. Зависимость распространения трех
близкородственных видов бокоплавов по течению реки
от концентрации солей в воде
(из Бигона, Харпера, Таунсенда, 1989).
Цифры внизу—расстояние от открытого моря.
Абиотические факторы оказывают существенное влияние на плотность популяций животных и растений. Понижение температуры часто катастрофически сказывается на популяциях животных: в районах, прилегающих
74















 к северным границам ареала, вид может стать редким и даже полностью исчезнуть. Кроме того, морозы в ряде случаев влияют и косвенно, поскольку пища, скрытая под толстым слоем льда или снега, становится совершенно недоступной для животных. В местах, подверженных сильным ветрам, рост растений задерживается, а фауна может быть частично или полностью уничтожена.
к северным границам ареала, вид может стать редким и даже полностью исчезнуть. Кроме того, морозы в ряде случаев влияют и косвенно, поскольку пища, скрытая под толстым слоем льда или снега, становится совершенно недоступной для животных. В местах, подверженных сильным ветрам, рост растений задерживается, а фауна может быть частично или полностью уничтожена.
ВИДОВОЙ УРОВЕНЬ
На уровне вида абиотические условия среды обитания нередко выступают как факторы, определяющие и ограничивающие географическое распространение. Кроме того, разнообразие климата, физических и химических свойств внешней среды в пределах обширных видовых ареалов обычно приводит к возникновению географической изменчивости видов. Основной способ адаптации видов к изменениям абиотических факторов — это приспособительная эволюция.
Экологические факторы обусловливают географическое распространение, т. е. влияют на положение, контуры и pазмеры ареалов. Эти показатели зависят от нескольких факторов одновременно, но среди них все же можно выделить основные. Для наземных видов таким фактором
чаще всего служит температура воздуха. Поскольку количество солнечного излучения, попадающегоназемную поверхность, растет в направлении от высоких широт к низким, ареалы большинства видов характеризуются широтной вытянутостью. Следует подчеркнуть, что границы ареалов видов часто определяются не средними,а крайними значениями температуры. Минимальная температура как ограничивающий фактор влияет на распространение многих теплолюбивых видов.
Перелетная саранча на севере Восточной Европы не заходит дальше июньской изотермы + 20 °С. В Скандинавии лось встречается на 10° севернее, чем в Сибири: несмотря на то, что средняя годовая температура воздуха в Сибири выше, чем в Скандинавии, зимы там гораздо более суровые. Северная
75
граница распространения змей на различных континентах в связи с температурными различиями последних неодинакова. В Европе она совпадает с 67° с. ш., в Азии — с 60°, в Америке—с 52° с. ш. Из растений в качестве примера можно привести марену Rubia peregrina, северная граница произрастания которой в Европе приблизительно определяется изотермой января + 4,5 °С. Ареал бука с севера ограничен изотермой -2°С.
Лимитирующим фактором для распространения холодостенотермных видов служит максимальная температура.
Для бабочки-капустницы, распространенной в Европе и Северо-Западной Африке, южной границей ареала является Палестина, поскольку там слишком высокие летние температуры. Ледниковые периоды позволили бореальным (северным) видам, не переносящим высоких температур, продвинуться далеко на юг, чуть ли не до Средиземноморского побережья. После отступания ледников эти виды остались в горных районах на территориях, значительно удаленных от их обычных ареалов,— так называемые бореально-альпийские виды. Хорошим примером такого вида служит бабочка торфяниковая желтушка, южная граница распространения которой в равнинных местах доходит до изотермы января - 2 °С, но которая обитает и на горных массивах к югу от основного ареала.
Температурные условия могут вызвать, наконец, географическую изменчивость видов. Проиллюстрировать это можно с помощью морфолого-зоогеографических закономерностей, к главнейшим из которых относится правило Бергмана. Немецкий физиолог Карл Бергман установил его еще в 1847 г. Теплокровным животным с постоянной и притом высокой температурой тела в жарком климате полезно излучать, а в холодном, наоборот, удерживать тепло. По мере же увеличения размеров животного поверхность тела возрастает в квадрате, а его объем — в кубе. Следовательно, поверхность крупного животного сравнительно меньше, чем у мелкого. Отсюда следует простой вывод, что теплокровным животным,
76
населяющим холодные страны, полезно быть крупными, а животным, населяющим теплые страны, — мелкими. Последующие наблюдения показали, что правило Бергмана соблюдается лишь в пределах подвидов одного вида или близких видов. При этом имеют значение не среднегодичные, а минимальные температуры, т. е. средние значения температур самого холодного месяца. Конкретным примером соблюдения правила Бергмана может служить изменение размеров бурого медведя: к северо-востоку Азии он становится все крупнее. Белый медведь крупнее бурого, а бурый — крупнее гималайского.
Второе морфолого-географическое правило, или правило Аллена, может быть сформулировано следующим образом: «У теплокровных животных по направлению к тропикам наблюдается удлинение хвостов, ушей, клювов, парных конечностей и различного вида выростов — хохлов, рожков, воротничков и т. д.». Это правило, как и другие морфолого-географические закономерности, приложимо лишь к подвидам одного вида или к близкородственным видам и может быть объяснено тем, что удлинение различного рода придатков тела способствует излучению тепла. Следовательно, правило Аллена представляет собой лишь частный случай правила Бергмана Проиллюстрируем справедливость правила Аллена несколькими примерами. Американская пустынная лисица, живущая в жарких пустынях Северной Америки, отличается от американской степной лисицы, распространенной в прериях, поджарым телом, длинными ногами и ушами. У обитающей в песчаных пустынях Северной Африки миниатюрной лисички из рода фенеков при высоте тела около 30 см уши достигают в длину 15 см и более.
Различие в значениях влажности в разных участках поверхности земли сказывается не только на характере распространения животных и растений, но и на окраске позвоночных животных. Об этом говорит правило Глогера — еще одна морфолого-географическая закономерность. Она выражается в том, что окраска позвоночных
77
животных, обитающих в странах с влажным климатом, как правило, темнее и ярче, чем у географических популяций тех же видов, распространенных в странах с сухим климатом. Иначе говоря, позвоночные животные, населяющие области с континентальным климатом, окрашены бледнее, чем родственные им формы в областях с морским климатом. Так, дальневосточный тигр заметно бледнее индийского; кавказского леопарда можно отличить от индо-африканского по более светлой окраске и т. д.
Особого внимания заслуживает желтовато-серая, так называемая пустынная, окраска, характерная для пустынно-степных животных, т. е. для обитателей областей с резко выраженным континентальным климатом. Приспособительное значение подобной окраски очевидно, так как делает ее обладателей незаметными на фоне песка (криптический эффект). Однако далеко не всем животным окраска с криптическим эффектом приносит пользу. Для летучих мышей, например, она не имеет большого значения, поскольку они днем прячутся в убежищах, кроме того, летящее животное обнаруживает себя уже благодаря резким движениям. Между тем у летучих мышей пустынных областей, в частности Средней Азии, мех и даже перепонки и когти имеют бледную серовато-желтую окраску. Возможно, такая окраска связана с какими-то физиологическими особенностями, определяющими возможность существования животных в области с пустынным климатом.

