




Основным способом питания растения является фотосинтез, в процессе которого под действием солнечной энергии происходит восстановление углекислого газа до углеводов (СН20):

80
где А-донор электронов. У зеленых растений (высшие растения, водоросли) донором электронов является вода, поэтому в результате фотосинтеза образуется кислород:
С02+Н20 -> (СН20) + 02.
У бактерий роль донора электронов могут выполнять, например, сероводород, органические вещества. Так, у зеленых и пурпурных серобактерий восстановление диоксида углерода происходит по схеме:
С02+ 2H2S -> (СН20) + 2S + Н20.
При фотосинтезе световая энергия улавливается хлоропластами и преобразуется в конечном итоге в энергию химических связей углеводов; в расчете на 1 грамм-атом поглощенного углерода фиксируется 114 ккал энергии. В процессе фотосинтеза участвуют как фотохимические реакции, так и чисто ферментативные (так называемые темновые) реакции и процессы диффузии, благодаря которым происходит обмен углекислотой и кислородом между растениями и атмосферным воздухом. Каждый из этих процессов находится под влиянием внутренних и внешних факторов и может ограничивать. продуктивность фотосинтеза в целом.
Растительная масса формируется не только за счет продуктов фотосинтеза. Наряду с углеродом, кислородом и водородом она содержит в среднем 2-4% азота (в белковых веществах – 15-19%). Среди биоэлементов азот по количеству в растениях занимает четвертое место. Между усвоением азота растением и продуктивностью существует корреляция; это относится как к отдельному растению, так и ко всему растительному покрову Земли. Прирост растительной массы нередко лимитируется количеством азота. При недостатке азота растения остаются низкорослыми, имеют мелкоклеточные ткани и грубые клеточные стенки.
Кроме светового питания растениям необходимо минеральное питание. Они нуждаются во многих элемен-
81
тах, которые либо поступают из минералов, либо становятся доступными в результате минерализации органического вещества. Все химические элементы поглощаются в форме ионов и включаются в растительную массу, накапливаясь в клеточном соке. После сжигания сухого органического материала минеральные вещества остаются в виде золы. В золе растений могут находиться все химические элементы, встречающиеся в литосфере. Жизненно необходимыми и незаменимыми являются основные элементы минерального пита ния, которые нужны в больших количествах: натрий, фосфор, сера, калий, кальций, магний, а также микроэлементы—железо, марганец, цинк, медь, молибден, бор и хлор. Кроме того, существуют элементы, которые требуются только для некоторых групп растений: натрий —для маревых, кобальт —для бобовых, алюминий—для папоротников и кремний—для диатомовых водорослей.
Для упорядоченного обмена веществ, хорошей продуктивности и беспрепятственного развития нужно, чтобы растение получало питательные вещества, включая микроэлементы, не только в достаточных количествах, но и в надлежащих соотношениях. Со времен Либиха известно, что урожай зависит от того вещества, которое имеется в недостаточном количестве. Разные виды растений значительно различаются по своим потребностям в питательных веществах. Для культурных растений этот вопрос изучен довольно хорошо. О специфических потребностях дикорастущих видов, напротив, известно немного, хотя именно эти сведения помогли бы лучше понять причины, определяющие характерный видовой состав сообществ.
Рассмотрим теперь роль питания в жизни животных. От наличия достаточного количества пищи зависят все формы их жизнедеятельности. Особенность действия пищи как экологического фактора для животных состоит в том, что экологический смысл имеет только нижний предел выносливости — в случае недостатка пищи она служит важным лимитирующим фактором, тогда как ее избыток не лимитирует развития особей (рис. 5.1). Как
82
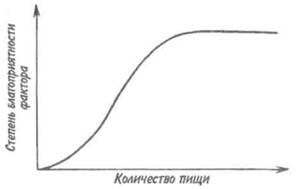
Рис. 5.1. Особенности действия пищи как экологического фактора в питании животных.
ограничивающий фактор недостаток пищи влияет на плодовитость и скорость развития животных.
У постельного клопа число отложенных яиц определяется количеством выпитой крови. Плодовитость колорадского жука увеличивается в зависимости от числа съеденных листьев. Точно так же развитие животных идет гораздо быстрее при достаточном количестве пищи. У жука-плавунца продолжительность третьей личиночной стадии равна 14 дням, если личинка получает ежедневно по 25 головастиков; если же она получает в день всего по одному головастику, эта стадия длится 51 день.
Необходимое количество корма возрастает с увеличением размеров животного. Однако, поскольку у мелких животных отношение площади поверхности тела к объему довольно велико, для них характерна более высокая интенсивность обмена и соответственно большая потребность в энергии на единицу массы тела, чем для крупных животных. Следовательно, мелким видам на единицу массы требуется больше пищи, чем крупным. Подобным же образом гомойотермные животные, которые должны поддерживать постоянную внутреннюю температуру тела, потребляют пищи больше, чем пойкилотермные.
Устойчивость к голоданию у пойкилотермных и гомойотермных животных также различна. У голодающих пойкилотермных животных с повышением температуры сре-
83
ды жизнь укорачивается (рис. 5.2, 2). Продолжительность жизни гомойотермных животных, подвергаемых голоду, тем больше, чем ближе температура среды к некоторому значению, которое ниже, но близко к температуре тела (рис. 5.2, 1).
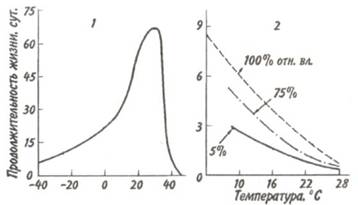
Рис. 5.2. Зависимость продолжительности жизни животных без пищи от температуры (из Дажо, 1975).
1 - домашний воробей, 2 Гусеница Panolis flammed.
Географическое распространение животных зачастую обусловлено пищевыми факторами. Например, бабочка грушевая плодожорка встречается только там, где произрастает дикая груша. Ареал желудевого долгоносика совпадает с областью распространения его кормового растения —дуба. Пальмовый орлан (Gypohierax angolensis) гнездится в Африке только там, где растет пальма Elalis guinensis, плодами которой он питается.
Наличие или отсутствие пищи существенно влияет на стациальное распределение животных: они предпочитают те биотопы, где есть подходящий для них корм. Нередко эта связь имеет очень жесткий характер. Например, многие растительноядные насекомые встречаются только на своих кормовых растениях.
Суточные, сезонные и иные регулярные миграции животных в большинстве случаев связаны с потребностями питания. Например, некоторые виды оленей но-
84
 чью пасутся на лугах, а днем перемещаются для отдыха в лес; летучие мыши, проводящие день в пещерах, ночью могут в поисках насекомых улетать от своих убежищ на десятки километров. Примеров сезонных миграций, вызванных изменениями количества корма, достаточно много. Усатые киты концентрируются в районах скопления зоопланктона, служащего им пищей. Ежегодные перелеты птиц с севера на юг также сформировались в эволюции как способ избежать зимней бескормицы.
чью пасутся на лугах, а днем перемещаются для отдыха в лес; летучие мыши, проводящие день в пещерах, ночью могут в поисках насекомых улетать от своих убежищ на десятки километров. Примеров сезонных миграций, вызванных изменениями количества корма, достаточно много. Усатые киты концентрируются в районах скопления зоопланктона, служащего им пищей. Ежегодные перелеты птиц с севера на юг также сформировались в эволюции как способ избежать зимней бескормицы.
Недостаточная обеспеченность кормом обычно вызывает у животных склонность к эмиграции из родных мест. Например, клесты-еловики в случае неурожая семян ели совершают далекие перелеты в поисках районов с обильным кормом. Для развития миграционного инстинкта необходимо, чтобы осенью в том районе, где обитает вид, пищевые ресурсы иссякли или стали недоступными, а в районе, куда вид перемещается, их было достаточно. Благодаря перелетам птицы наводняют биотопы, не полностью используемые оседлыми видами. Другими словами, перелетные виды используют сезонные кормовые излишки.
С пищей животные и растения получают, во-первых, энергию, необходимую для жизнедеятельности, и, во-вторых, питательные вещества, необходимые для построения тканей тела и выполнения всех физиологических функций. Требования к пище могут значительно меняться в зависимости от состояния организма, времени года и т. п. Для животных и растений имеют значение как количество питательных веществ, так и их качественный состав.

