




Грибы - одно из самых больших царств организмов. Появились первые представители 450-500 млн. лет назад. Их группы произошли независимо, от разных жгутиковых. Некоторые предположительно произошли от водорослей или амебоидных предков. Существует около 120000 видов грибов, разнообразных по внешнему виду и распространению.
По наличию в обмене веществ мочевины, хитина в клеточной стенке клеток, запасного продукта – гликогена грибы приближаются к животным. А по способу питания путем всасывания (абсорбтивное питание), а не заглатывания пищи, и по неограниченному росту они напоминают растения.
Тело гриба представляет собой мицелий (грибницу), который состоит из разветвленной сети тонких нитей – гиф. У грибов, условно называемых низшими, мицелий неклеточный (не имеет перегородок) и представляет собой одну разветвленную клетку; у остальных - гифы разделены перегородками (септами) на отдельные клетки, образуя многоклеточный мицелий. В септах многоклеточного мицелия имеются отверстия (поры), через которые осуществляется цитоплазматическая связь между клетками. Кроме того, гифы мицелия могут сильно переплетаться, образуя ложную ткань – плектенхиму. У некоторых грибов – паразитов мицелий отсутствует, а тело представляет собой плазмодий.
Клетки имеют клеточную стенку и протопласт. Клеточная стенка у низших грибов состоит из пектиновых веществ, у высших – из углеводов близких к целлюлозе, со значительной примесью хитина (близок к хитину насекомых). В протопласте выделяется цитоплазма (у молодых клеток гомогенна), в которой хорошо различима плазмалемма и тонопласт, рибосомы, митохондрии, аппарат Гольджи, эндоплазматическая сеть, 1-2 или много мелких ядер. Пластид нет. Запасные вещества – гликоген, жиры, волютин, гранулы белков (в вакуолях), крахмала нет.
Грибы – гетеротрофы (питаются готовыми органическими веществами). По образу жизни их делят на две группы. Первая – сапротрофы (получают органические вещества из органических останков), вторая – паразиты (используют живую субстанциюрастений, животных, человека и др. в качестве источника органических веществ). К первой группе относится большинство грибов. Питание грибов происходит путем осмоса всей поверхностью мицелия, реже при помощи всасывающих ризоидов (плесневые грибы) или присосок – гаусторий у паразитов (ржавчинные, головневые грибы).
Размножаются грибы вегетативным, бесполым и половым путем. Вегетативное размножение грибов происходит: 1) частями мицелия; 2) оидиями (артроспорами) - образуются в результате распадения гиф на отдельные клетки, дающие новый мицелий; 3) хламидоспорами ( образуются как оидии) - имеют более твердую, часто темноокрашенную оболочку (головневые грибы) и предназначены для перенесения неблагоприятных условий; 4) почкованием - на мицелии образуются боковые выросты, которые отчленяются от него.
Бесполое размножение у грибов происходит тремя способами: зооспорами, спорангиоспорами и конидиями. Зооспоры (преимущественно у водных грибов) имеют жгутики, подвижны и образуются в зооспорангиях. Спорангиоспоры – это неподвижные споры, которые развиваются в спорангиях. Конидии характерны для высших грибов (аскомикоты, базидиомикоты, дейтеромикоты) и образуются на особых гифах – конидиеносцах. При этом кончик конидиеносца отделяется перегородкой, округляется и отпадает в виде конидии. Под первой конидией до ее отделения таким же образом формируется следующая и т.д., в результате образуется целая цепочка.
Половое размножение у грибов очень разнообразно и отмечено у всех таксонов, кроме дейтеромикотов (рис. 85). Причем половой процесс состоит из двух этапов: 1 - слияние цитоплазмы двух клеток (плазмогамия); 2 - слияние ядер (кариогамия); и эти этапы разделены во времени.

Рисунок 85 - Некоторые типы полового процесса у грибов
Типы полового размножения:
гаметогамия – слияниие гамет, образующихся в гаметангиях;
гаметангиогамия - слияние содержимого двух многоядерных специализированных половых органов (мужского и женского гаметангиев), которые не дифференцированы на гаметы; в качестве гамет здесь можно рассматривать свободно располагающиеся в гаметангиях ядра; характерен для зигомикотов и аскомикотов;
соматогамия - половых органов не образуется, а сливаются гаплоидные соматические клетки физиологически различных гиф; характерен для базидиомикотов;
хологамия (у грибов с одноклеточным мицелием) - наблюдается слияние двух особей.
Половой процесс у так называемых высших грибов завершается образованием зиготы, ее мейотическим делением и образования полового спороношения.
В систематике грибов еще очень много спорного. До недавнего времени существовало деление грибов на низшие (хитридиомицеты, оомицеты и зигомицеты) и высшие (аскомицеты, базидиомицеты и дейтеромицеты). В настоящее время, к настоящим грибам относят четыре таксона, которые имеют общее происхождение – зигомикоты, аскомикоты, базидиомикоты, дейтеромикоты (несовершенные грибы) и лишайники, слоевища которых образованы мицелием грибов. Оомикоты и Хитридиомикоты отнесены к грибоподобным протоктистам. Перечисленные таксоны обычно считают классами (имеют окончания - мицета, - mycetes) или отделами (имеют окончание - микота, - mycota).
2 Отдел Слизевики, или Миксомицеты.
Отдел объединяет бесхлорофилльные организмы, которые по своему строению и образу жизни занимают промежуточное положение между животными и грибами. Одни из них имеют микроскопически малые размеры и представлены одно- или многоядерной амебоидной клеткой, другие крупные, многоядерные, обычно подвижные (0,1-0,4 мм/мин), бесцветные или окрашенные, подчас достигающие более 30 см. В состав плазмодия входят белки, гликоген, жиры, пигменты и другие вещества.
Вегетативное размножение у большинства слизевиков происходит путем деления амебоидных клеток. У ряда сапротрофных форм содержимое плазмодия идет на образование плодовых тел – спорангиев, внутри которых возникают одноклеточные споры с плотной целлюлозной оболочкой. У паразитов спорангии не образуются и плазмодий распадается на споры в клетках хозяина, служащих им вместилищем.
Зрелые споры на влажном субстрате прорастают зооспорами с двумя гладкими жгутиками неравной длины, а в случае низкой влажности среды формируются миксоамебы (амебы). Зооспоры и миксоамебы могут размножаться делением. При определенной концентрации клеток они попарно сливаются, образуя диплоидные клетки, из которых в дальнейшем развивается плазмодий. У миксомицетов с псевдоплазмодиями половой процесс отсутствует, а псевдоплазмодий возникает путем агрегации миксоамеб.
Миксомицеты – преимущественно сапротрофы, обитают на отмершей древесине, коре деревьев, лесной подстилке и аналогичных субстратах; паразитные формы развиваются в клетках водорослей, грибов и высших растений.
Отдел Миксомикота объединяет три класса: Протостелиомицеты (Protosteliomycetes), Миксогастеромицеты (Myxogasteromycetes), и Цератиомиксомицеты (Сeratiomyxomycetes). В основу такого деления положены степень организации тела и особенности циклов развития представителей разных классов.
Класс Миксомицеты (Myxomycetes) или Миксогастеромицеты, (Myxogasteromycetes)
К этому классу относится подавляющее большинство миксомицетов со всеми характерными признаками отдела. Свободноживущий плазмодий проявляет положительные гидро- и трофотаксисы и отрицательный фототаксис. За счет циркуляции цитоплазмы он активно передвигается, образуя псевдоподии, которыми захватывает пищу – бактерий, амеб, мелких беспозвоночных животных, различные твердые пищевые частицы. К концу вегетации плазмодии заметно увеличиваются в размерах и выползают на поверхность субстрата, т. е. отрицательный фототаксис сменяется положительным. Здесь плазмодии уплотняются и превращаются в плодовые тела различной формы, размеров, окраски, на ножках или сидячие. У многих миксомицетов в плодовых телах кроме спор имеются особые нити - капиллиций, который разрыхляет споровую массу и способствует рассеиванию спор. Вид и строение капиллиция являются характерными систематическими признаками рода или вида. У некоторых представителей образуется псевдокапиллиций, представляющий собой уплощенные ветвящиеся образования с поперечными складками на поверхности и формирующийся из остатков стенок спорангия.
При благоприятных условиях спора прорастает 1–4 зооспорами или миксоамебами, способными в капельножидкой среде вырабатывать жгутики и превращаться в зооспоры. Затем зооспоры или миксоамебы попарно копулируют, образуя диплоидную миксоамебу, из которой возникает плазмодий.
Миксогастеромицеты отличаются поразительной способностью приспосабливаться к неблагоприятным условиям среды (отсутствие пищи, большие морозы, низкие значения рН и др.). Их плазмодии могут превращаться в утолщенную твердеющую массу – склероции, которые сохраняют жизнеспособность в течение многих лет. Часть зооспор и миксоамеб при экстремальных условиях могут превращаться в цисты.
Класс объединяет более 400 видов, относящихся к 4 порядкам: лициальные, трихиальные, физаральные и стемонитальные. В основе деления на порядки лежат особенности строения плазмодия, спорангиев, наличие настоящего и ложного (псевдокапиллиция) капиллиция:
Порядок Лициальные (Liciales). У лициальных спорангии шаровидные или в виде плазмодиекарпиев, покрытых перепончатой или хрящеватой оболочкой (перидием), без настоящего капиллиция.
У рода ликогала (Lycogala) плазмодий имеет кораллово-красную окраску. Эталии округлые, диаметром 0,3–1,5 см, сидячие, скученные или разбросанные на поверхности субстрата. Вначале они имеют ту же окраску, что и плазмодий, позже буреют. При их созревании перидий становится тонким, бородавчатым, с отверстием сверху для выхода спор. В таком виде эталии ликогалы похожи на мелкие грибы-дождевики. Псевдокапиллиций представлен сеткой из трубчатых волокон с булавовидными концами. Споровый порошок розовый или оранжевый.
Сапротрофы на отмершей древесине и других субстратах. Известно 4 вида, распространенных в умеренных поясах земного шара. В Беларуси встречается Л. древесинная, или «волчье вымя» (L.epidendrum).
Порядок Трихиальные (Trichiales). У трихиальных плазмодий бесцветный, беловатый, розоватый, красный или пурпурово-коричневый. Спорангии овальные, сидячие или на коротких ножках, одиночные или чаще скученные. У всех видов есть настоящий капиллиций, характерный для каждого рода.
Род трихия (Trichia) является наиболее типичным для порядка. У его представителей спорангии групповые, цилиндрические или бокаловидные, сидячие или на ножках Перидий гладкий, блестящий или матовый, часто с концентрическими утолщениями или бородавками. Капиллиций образован свободными, простыми или ветвящимися полыми внутри, заостренными на концах нитями с лентовидными спиральными утолщениями. Споры шаровидные с сетчатой или шиповатой поверхностью, в массе желтые, оранжево-желтые или красноватые.
Известно 12 видов, распространенных на всех континентах. В Беларуси они встречаются в основном на отмершей древесине. Наиболее часто встречаются Т. изменчивая (Т. varia), Т. гроздевидная (Т. botrytis), Т. обманчивая (Т. decipiens).
Порядок Физаральные (Physarales). Плазмодий сильно гранулированный, бесцветный или яркоокрашенный. Спороношение в виде эталиев или одиночных, сидячих или на ножках спорангиев. Капиллиций чаще состоит из ветвящихся нитей, образующих узловые расширения. Для видов этого порядка характерны большие скопления извести, главным образом на поверхности перидия, ножках спороношения и капиллиций. Споры в массе своей темно-пурпурные, фиолетово-коричневые, от темно-коричневых до черных.
Род физарум (Physarum) содержит виды с белым, сероватым, желтым или оранжевым плазмодием. Спорангии округлые, сидячие или на ножках, иногда в виде продолговатых изогнутых плазмодиекарпиев. Перидий, как правило, инкрустирован известью, хрупкий, неправильно растрескивающийся при созревании. Капиллиций в виде сети тонких разветвленных нитей, прикрепленных к основанию спорангия или к различным участкам перидия, с многочисленными расширениями и вздутиями. Споры в массе от фиолетовых до черных.
Известно более 70 видов, распространенных по всему земному шару. В Беларуси их 6. Сапротрофы. Развиваются в июле–сентябре на отмершей древесине, лесной подстилке, а также на плодовых телах макромицетов. Наиболее распространены 2 вида: Ф. пепельный (Ph. cinereum), часто образующий спороношения даже на живых растениях, и Ф. поникший (Ph. nutans), развивающийся лишь на отмершей древесине, мхе, сухой соломе.
Род фулиго (Fuligo) включает виды, у которых плазмодий ярко-желтый, реже белый или кремовый, достигающий иногда нескольких десятков сантиметров. Эталии подушковидные, сильно варьируют в размерах, форме и окраске: белые, охряные, зеленоватые, розоватые, тускло-красные, почти коричневые. Толстый, хрупкий перидий легко разрушается и отслаивается кусками, обнажая почти черную массу спор. Капиллиций состоит из очень тонких, бесцветных анастомозирующих нитей, соединенных вздутиями (узлами), на которых расположены бесцветные или желтоватые гранулы извести.
В роде более 10 видов, распространенных во многих странах мира. Наиболее известен Ф. гнилостный (F. septica), развивающийся в июле–октябре на гнилых пнях, коре ветвей деревьев, засохших листьях в оранжереях.
Порядок Стемонитальные (Stemonitaies). Он объединяет миксомицеты с тонким прозрачным плазмодием. Спороношение в виде отдельных спорангиев или эталиев, покрытых пленчатым, быстро исчезающим перидием.
Род стемонитис (Stemonitis) – типичный представитель порядка. У видов рода плазмодий сначала бесцветный, а затем беловатый или лимонно-желтый. Спорангии цилиндрические, высотой 2–20 мм, на длинных ножках, которые внутри спорангия превращаются в колонку. Перидий вскоре исчезает, обнажая капиллиций в виде ажурной сеточки вокруг колонки. Капиллиций одного цвета с колонкой и ножкой, покрыт коричневым, буровато-фиолетовым или почти черным споровым порошком.
Класс Аскимицеты.
Аскомицеты, или сумчатые грибы, - один из обширнейших классов грибов, включающий около 30 тыс. видов, что составляет около 30% всех известных видов грибов.
Основной признак аскомицетов - формирование в результате полового процесса сумок, или асков, - замкнутых одноклеточных структур, содержащих определенное число аскоспор, обычно восемь.
Вегетативное тело аскомицетов - разветвленный гаплоидный мицелий, состоящий из одноядерных или многоядерных клеток. У некоторых низших аскомицетов (дрожжи) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися или делящимися клетками. Основные полисахариды, входящие в состав клеточных стенок аскомицетов - хитин и глюканы.
В цикле развития многих аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения - конидии. Конидиальные спороношения развиваются в период вегетации грибов и служат для их массового расселения.
Половой процесс, типичный для аскомицетов, - гаметангиогамия, т.е. слияние двух гаметангиев - специализированных клеток, не дифференцированных на гаметы.
У низших аскомицетов (подкласс голосумчатые) половой процесс сходен с зигогамией у зигомицетов, но при этом сливаются только два ядра и зигота развивается в сумку. Высшие аскомицеты имеют довольно сложные гаметангии. Женский гаметангий состоит из двух частей - аскогона и нитевидно вытянутой трихогины, мужской гаметангий - антеридий - одноклеточный. При слиянии содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола сразу не сливаются, а объединяются попарно, образуя дикарион. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. На аскогенных гифах развиваются сумки. Сумка увеличивается в размерах, ядра дикариона сливаются, диплоидное ядро делится редукционно и митотически и вокруг восьми гаплоидных ядер формируются аскоспоры. В сумке аскоспоры окружены неиспользованной на их формирование цитоплазмой - эпиплазмой. К моменту созревания аскоспор в цитоплазме происходит превращение гликогена в сахар, тургорное давление в сумке резко возрастает и аскоспоры с силой выбрасываются.
В результате образования аскогенных гиф увеличивается число сумок, а следовательно, и аскоспор, развивающихся из одного аскогона.
В цикле развития аскомицетов чередуются три фазы: длительная - гаплоидная, в течение которой происходит бесполое размножение, непродолжительная - дикариотическая (аскогенные гифы) и очень короткая - диплоидная (молодая сумка с ядром).
Аскомицеты широко распространены во всех географических зонах. Среди них есть как сапротрофы, так и паразиты. Аскомицеты вызывают заболевания многих культурных растений, вместе с тем они используются как продуценты антибиотиков, витаминов, ферментов и алколоидов, а также как возбудители спиртового брожения.
Аскомицеты отдел в царстве грибов, объединяющий организмы с септированным (разделённым на части) мицелием и специфическими органами полового спороношения - сумками (асками), содержащими чаще всего по 8 аскоспор. Имеют и бесполое спороношение, причём во многих случаях половой процесс утрачивается, и такие виды грибов относят к несовершенным грибам.Сумки могут образовываться либо непосредственно на мицелии (у низших аскомицетов), либо в специальных вместилищах - плодовых телах и аскостромах(у высших аскомицетов). На основании отсутствия или наличия плодовых тел класс Аскомицеты делят на три подкласса. Наиболее важные из них: 1) подкласс гемиаскомицеты, или голосумчатые (Hemiascomycetidae) и 2) подкласс эуаскомицеты (Euascomycetidae).
У сумчатых грибов имеются четыре типа плодовых тел:
Клейстотеции (клейстокарпии) представляют собой полностью замкнутое плодовое тело с находящимися внутри асками, освобождающимися после разрушения его стенок. Характерны для группы порядков плектомицеты.
Перитеции (др.-греч. περι- возле, около, θήκη - хранилище) — почти замкнуты («полузамкнуты»), то есть сумки окружены перидием, обычно имеют кувшинообразную форму с выводным отверстием в верхней части. Характерны для группы порядков пиреномицеты.
Апотеции - открытые вместилища аск. Образуют чаши (блюдца). По верхней стороне плодового тела расположен слой сумок и парафиз; парафизы, пока сумки незрелые, могут смыкаться над ними своими вершинами и выполнять защитную функцию. Также образуют сморщенную поверхность сморчков (причём рёбра складок стерильны), могут, однако, не раскрываться (у трюфелей). Аскомицеты с апотециями считаются наиболее высокоорганизованными и объединяются в группу порядков дискомицеты.
Псевдотеции характерны для класса Loculoascomycetes. При их формировании вначале образуются мицелиальные стромы, в полостях которых формируются гаметангии, происходит половой процесс и образование сумок. Клейстотеции и перитеции часто развиваются на особых образованиях - стромах - плотных сплетениях гиф, которые часто ошибочно принимают за плодовые тела. Они могут находиться на поверхности стромы или быть погружёнными в её тело. Представитель: Saccharomycetes - Дрожжи.
Класс Голосумчатые грибы (Hemiascomycetes). Сумки развиваются непосредственно на мицелии, плодовых тел нет. Так виды рода Saccharomyces (дрожжевые грибы) представляют собой одиночные, одноядерные, сферические или овальные клетки, которые размножаются вегетативно почкованием. Последнему способствует достаточное количество сахара, температура – 25-30°С и аэрация. Только в результате интенсивного размножения возникают цепочки клеток (псевдомицелий), которые в последствие распадаются (рис. 86).

1 – клетка дрожжей, 2 – начало почкования, 3 – почкование (образование псевдомицелия), 4 – сумка со спорами
Рисунок 86 - Хлебные дрожжи (Saccharomyces cerevisiae)
При истощении питательной среды у дрожжей можно наблюдать половой процесс. Он состоит в слиянии (копуляции) двух гаплоидных клеток. Впоследствии из зиготы образуется сумка с 4-8 аскоспорами. Рассматриваемый род объединяет как природные виды, так и «культурные» (производственные), существование которых тесно связано со всей историей бродильной промышленности. Большое практическое значение имеют пивные, или хлебные дрожжи (Saccharomyces cerevisiae), которые существуют только в виде культурных рас и винные дрожжи (S. vini, S. ellipsoideus), которые живут на поверхности и в соке различных плодов.
Другие представители данного класса (например виды рода Taphrina – тафрина), вызывают поражение стеблей, листьев, цветков, плодов растений, способствуют их разрастанию и деформации.
Класс Плодосумчатые грибы (Carpoascomycetes). У представителей этого класса сумки развиваются в специальных вместилищах – плодовых телах, различных по форме и величине. Существует несколько типов плодовых тел: клейстотеций (замкнутое, шарообразное плодовое тело), перитеций (полузамкнутое, кувшиновидное плодовое тело с отверстием на вершине) и апотеций (открытое, чашевидное плодовое тело).
Представитель данного класса - Спорынья (Claviceps) паразитирует на злаках (рожь), осоках. Имеет плодовое тело перитеций. В конце лета в колосьях ржи вместо зерновок образуются черно-фиолетовые рожки (склероции), которые состоят из плотного переплетения гиф.
Цикл развития спорыньи включает три стадии: склероциальную, сумчатую и конидиальную.
Стадия I - образование склероция (покоящейся стадии гриба). При сильном поражении ржи на отдельных колосьях может быть до 3-4 склероциев. Далее при уборке хлеба склероции могут самопроизвольно опадать на землю (они хорошо переносят морозы и на следующий год после всходов ржи начинают прорастать), или при обмолоте попадать в товарное или семенное зерно.
Стадия II - на прорастающем склероции появляются красные или темно-розовые булавовидные плодовые тела, состоящие из тонких ножек и шаровидных головок, усаженных многочисленными мелкими коническими выступами ("бородавочками"). Эта стадия – строма. Бородавочки на головке являются выходами перитециев - яйцевидных полостей, образующихся в периферической части головки. В перитециях вырастают многочисленные булавовидной формы сумки, в каждой из которых развивается по 8 нитевидных аскоспор. К моменту цветения ржи плодовые тела гриба полностью созревают; при этом из слизисто разбухающих перитециев выдавливаются сумки, которые лопаются; при этом из них выбрасываются аскоспоры и воздухом разносятся по цветущей ржи.
Стадия III начинается с попадания аскоспор на перистые рыльца цветков ржи и их прорастания. Из сплетения гиф на завязи цветка образуется грибница, по мере развития которой начинается бесполое размножение гриба. Заключается оно в отшнуровывании с концов гиф многочисленных мелких эллиптических конидиоспор. Одновременно грибницей вырабатывается клейкая жидкость, содержащая сахаристые вещества, называемая "медвяной росой". Капли последней стекают по пораженному колосу, унося с собой конидиоспоры. Сладкая жидкость привлекает насекомых, которые, перелетая на другие колосья, разносят конидиоспоры, способствуя, тем самым, новому (повторному) заражению ржи. Конидиоспоры, попав на здоровые цветки ржи, также прорастают, образуя на завязи грибницу. Постепенно грибницы (образовавшиеся как из аскоспор, так и из конидиоспор), разрастаясь, разрушают завязь, и, в конечном счете, на месте и вместо зерна развивается белое продолговатое крупное грибное тело - молодой склероций. К моменту созревания ржи созревают и склероции; гифы уплотняются, наружный слой склероция при этом пигментируется, окрашиваясь в темно-фиолетовый цвет (рис. 87).
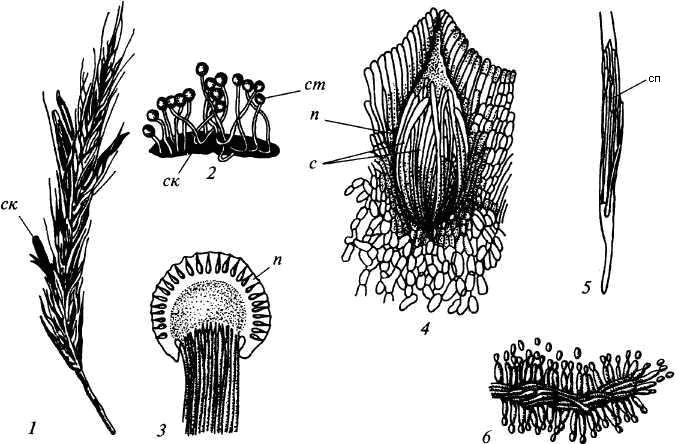
1 – колос ржи со склероциями, 2 – склероций, проросший головчатыми стромами, 3 - разрез стромы с перитециями, 4 – отдельный перитеций в строме с сумками, 5 – сумка с аскоспорами, 6 – конидиальное спороношение, ск – склероции, ст – стромы, п – перитеций, с – сумки, сп – споры
Рисунок 87 – Цикл развития с порыньи пурпурной (Clavicepspurpurea)
Рожки (склероции) спорыньи эрготаминового (эрготоксинового) штамма (Cornua Secalis cornuti stamm Ergotamini (Ergotoxini)) содержат алкалоиды индольного ряда, обладающие ядовитыми свойствами и оказывающие сложное влияние на организм человека. В современной медицине алкалоиды спорыньи широко применяются для лечения сердечно-сосудистых и нервных заболеваний (адреноблокирующая активность), а также в акушерской практике (вызывают сокращение матки).
Плодовое тело - апотеций имеют пецица (Peziza), строчок (Gyromitra), сморчок (Morchella) (рис. 88).

Рисунок 88 - Апотеции пецицевых: 1 – пецица, 2 - сморчок, 3 – строчок
Пецица встречается в лесах, на местах пожарищ, на почве огородов, коровьем навозе. Строчок и сморчок встречаются в лесу рано весной. Плодовое тело состоит из шляпки с морщинистой поверхностью, выстланной гимением (слой асков, разделенных парафизами (бесплодными гифами) и ножки. Строчки содержат ядовитую гельвелловую кислоту (разрушается после длительного кипячения).
Основная литература:
1 Еленевский А.Г., Соловьев М.П., Тихомиров В.Н. Ботаника: систематика высших, или наземных, растений. 2 изд. - М.: Academіa, 2001. - 429 с.
2 Нестерова С.Г. Лабораторный практикум по систематике растений. - Алматы: Қазақ ун-ті, 2011. - 220 с.
3 Родман А.С. Ботаника. – М.: Колос, 2001. - 328 с.
Дополнительная литература:
1 Абдрахманов О.А. Систематика низших растений. – Караганда: Изд-во КарГУ, 2009. - 188 с.
2 Билич Г.Л., Крыжановский В.А. Биология. Т. 2: Ботаника. - М.: Оникс 21 век, 2002. - 543 с.
3 Абдрахманов О.А. Практические работы по систематике низших растений. Ч. 2. Грибы и водоросли. – Караганда: Изд-во КарГУ, 2001. - 144 с.
4 Абдрахманов О.А. Лабораторный практикум по бактериям и водорослям. Учебное пособие. - Алматы: Казакадем образование, 2000. - 130 с.
5 Абдрахманов О.А. Практические работы по систематике низших растений. Ч. 2. Грибы и водоросли. – Караганда: Изд-во КарГУ, 2001. - 144 с.
Контрольные вопросы:
1 В чем отличие грибов от друг групп организмов?
2 Опишите циклы размножения и смены фаз у слизневиков.
3 Какие жизненные формы представлены у аскомицетов?
4 Опишите строение клетки гриба и слизневика.
5 Какое значение имеют слизневики и грибы в экологии и в жизни человека?
6 Какие запасные питательные вещества образуют грибы?
7 Опишите сходные и отличительные черты строения слизневиков и аскомицетов.

