




Отдел насчитывает около 1200 видов из 120 родов, объединяющих эукариотические одноклеточные (в том числе - двужгутиковые), коккоидные и нитчатые формы. Группа совмещает в себе признаки растений и животных: у некоторых видов имеются щупальца, псевдоподии и стрекательные клетки; некоторые имеют характерный для животных тип питания, обеспечивающийся глоткой. У многих имеется стигма, или глазок. Клетки часто покрыты твердой оболочкой. Хроматофоры буроватых и красноватых оттенков, содержат хлорофиллы а и с, а также каротины, ксантофиллы (иногда - фикоцианин и фикоэритрин). В качестве запасных веществ откладывается крахмал, иногда - масло. На поверхности клетки и в глотке имеются бороздки (рис. 72).

Рисунок 72 – Некоторые пиррофитовые водоросли
Наиболее существенным, общим для всех пирофитовых водорослей признаком является спинно-брюшное (дорсовентральное) строение их клеток. У громадного большинства представителей этого отдела в строении клеток отчетливо выражены спинная, брюшная и боковые стороны, а также хорошо заметна разница между передним и задним концами.
Наличие у клеток бороздок является вторым важным признаком для всех пирофитовых. Бороздок может быть две: продольная и поперечная – или одна продольная. Располагается продольная бороздка всегда на брюшной стороне клетки. У представителей с двумя бороздками место их пересечения такжеприурочено к брюшной стороне клетки.
Два разных по длине, строению и даже функциям жгутика – третий общий признак для всехподвижных одноклеточных (монадных) форм пирофитовых. Эти жгутики получили разные названия: плавательный и рулевой – у хлоромонадофитовых, передний и задний – у криптофитовых, поперечный и продольный – у динофитовых.
Другой очень важный отличительной чертой является присутствие у громадного большинства одноклеточных пирофитовых так называемой глотки, обычно имеющей вид мешка, трубы, внутреннего кармана или треугольного резервуара, а также наличие особых, сильно преломляющих свет телец - трихоцист. Последние имеют вид одиночных или многочисленных булавовидных или игловидных образований, располагающихся или в периферическом слое цитоплазмы (у хлоромонадофитовых), илипродольными рядами по внутренней поверхности глотки (у криптофитовых), или внутри протопласта (удинофитовых).
Особо следует сказать о хлоропластах. По разнообразию их окраски пирофитовым принадлежитпервое место среди водорослей. Обычно хлоропласты пирофитовых окрашены в оливковый, бурый или коричневый цвет, нередко наблюдаются желтый, золотистый, красный, реже голубой и синий, а также всевозможные оттенки указанных цветов. Лишь чисто зеленая окраска хлоропластов у них полностьюотсутствует. Как и в составе других отделов, здесь имеются также и неокрашенные формы.
Питание – сапрофитное или автотрофное, иногда наблюдается смешанный тип питания. Размножаются делением в подвижном или неподвижном состоянии (вегетативно), зооспорами и автоспорами. Половое размножение известно у немногих форм; оно проходит в виде слияния изогамет.
Пирофитовые водоросли - обычные обитатели загрязненных водоемов: прудов, отстойников, некоторых водохранилищ и озер. Многие образуют фитопланктон в морях. При неблагоприятных условиях образуют цисты с толстыми целлюлозными оболочками.
Практическое значение пирофитовых водорослей заключается прежде всего в их активном участии в круговороте веществ в водоемах. Они способны давать большую биомассу и поэтому важнымзвеном в цепи питания рыб и других гидробионтов, а также играют существенную роль в отложении сапропелей.
Будучи весьма чувствительными к органическим загрязнениям в воде или, наоборот, требуя для своегоразвития определенного содержания органического вещества в окружающей среде, многие виды пирофитовых водорослей используются в качестве показательных форм сапробности при биологическом анализе воды, т.е. являются так называемыми биоиндикаторами при санитарно-биологической оценке вод. Многие пирофитовые, обитающие в загрязненных и сточных водах, выполняют функцию активных санитаров этих вод, так как принимают участие в процессах их самоочищения. Среди пирофитовыхизвестны и ядовитые формы. Наконец, панцирные формы благодаря окремнению и обызвествлению панциря хорошо сохраняются в осадочных образованиях и, подобно диатомовым, широко используются вгеологии и практике поисковых работ для определения возраста осадочных пород (перидинеи из подотделадинофитовых).
Наиболее широко распространен и богат видами род Криптомонада (Cryptomonas).
Золотистые водоросли.
Отдел Золотистые водоросли (Chrysophyta). Микроскопические или мелкие (до 2 см длины) организмы золотисто-желтого цвета, обитающие в соленых и пресных водоемах по всему земному шару. Имеются одноклеточные, колониальные и многоклеточные формы (рис. 73).
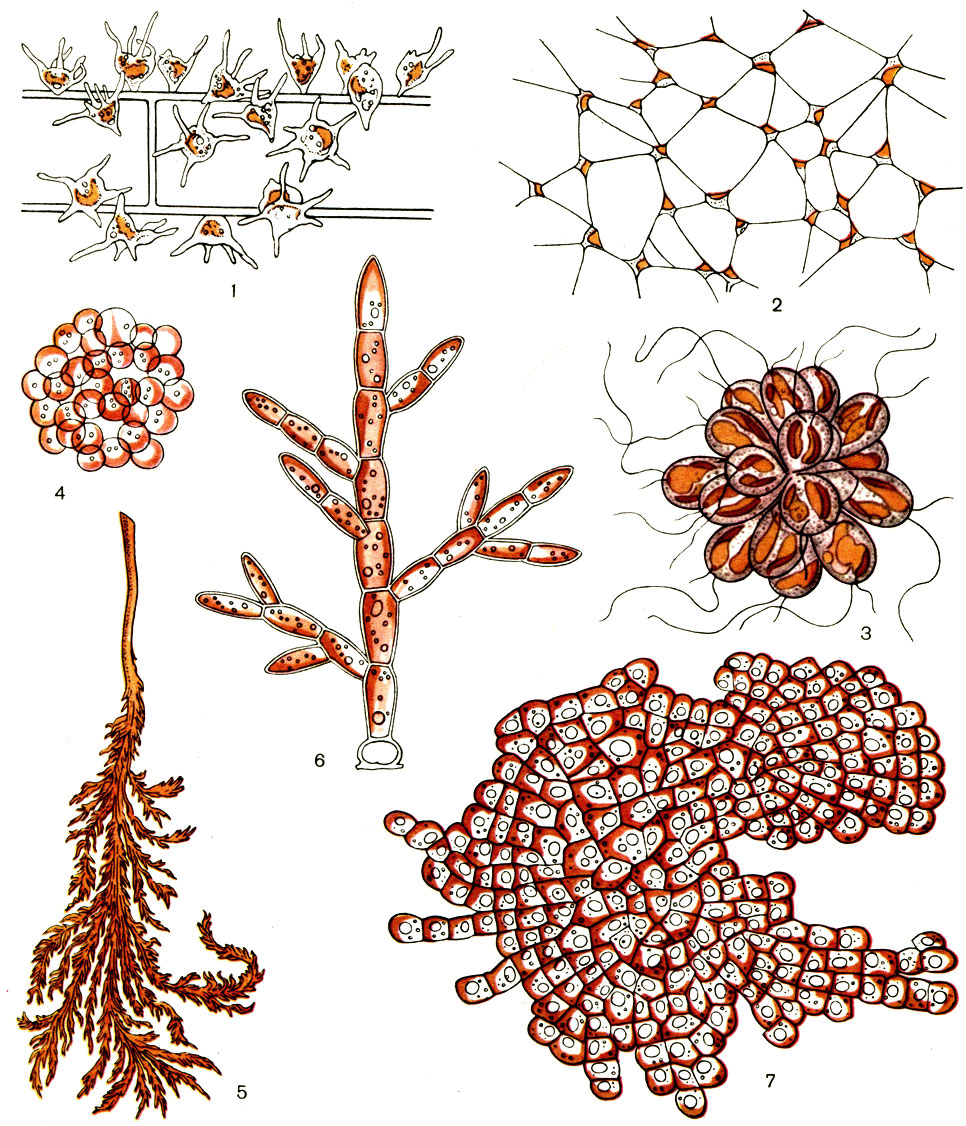
Рисунок 73 – Золотистые водоросли
Отличаются значительным морфологическим разнообразием. Золотистые водоросли (фото их представлено выше) бывают как одноклеточными, так и многоклеточными, колониальными. Кроме того, среди золотистых водорослей есть весьма своеобразный представитель. Многоядерный таллом у него является голым плазмодием. Таким образом, весьма разнообразны золотистые водоросли. Строение клетки этих организмов характеризуется наличием разного количества жгутиков. Их число зависит от вида. Обычно их два, однако нужно отметить, что у некоторых видов золотистых водорослей имеется три жгутика. Третий, неподвижный, расположен между двумя подвижными. Он называется гантонемой и характеризуется расширением на конце. Функция гантонемы заключается в том, что клетка с ее помощью прикрепляется к субстрату.
Золотистые водоросли – это отдел, который включает в себя преимущественно микроскопические виды. Их хлоропласты, как правило, золотисто-желтого цвета. Из пигментов следует отметить хлорофилл А. Кроме того, был обнаружен хлорофилл Е, а также множество каротиноидов, включая каротин и ряд ксантофиллов, в основном золотистый фукоксантин. В качестве запасных веществ откладывается хризоламинарин и масло. Некоторые виды гетеротрофны.
Окраска представителей интересующего нас отдела может иметь разные оттенки, в зависимости от преобладания того или иного из этих пигментов. Она может быть от зеленовато-бурой и зеленовато-желтой до чисто золотисто-желтой.
Золотистые водоросли, виды которых многочисленны, – это фототрофные организмы. Их значение заключается, главным образом, в создании в водоемах первичной продукции. Кроме того, они участвуют в цепи питания различных гидробионтов, включая рыб, золотистые водоросли. Виды их улучшают газовый режим различных водоемов, где они произрастают. Также они формируют отложения сапропеля. Отдел Золотистые водоросли характеризуется размножением его представителей с помощью простого клеточного деления, а также с помощью распада многоклеточного таллома или колоний на отдельные части. Ученым известен и половой процесс, представляющий собой типичную автогамию, хологамию или изогамию. В результате процесса размножения появляются эндогенные кремнистые цисты, различающиеся по такому основанию, как характер скульптурированности их оболочки. Эти цисты выполняют важную функцию – они помогают водорослям пережить неблагоприятные условия.
Большинство форм имеют 1-2 жгутика и в силу этого подвижны. Размножаются преимущественно бесполым путем - делением или зооспорами; половой процесс известен лишь у нескольких видов. Встречаются обычно в чистых пресных водах (кислые воды сфагновых болот), реже - в морях и в почвах. Типичные элементы фитопланктона.
Отдел Золотистые водоросли включает в себя представителей нескольких классов. Ниже мы кратко охарактеризуем некоторые из них.
Класс Хризокапсовые. Его представители отличаются наличием сложного таллома, который представлен слизистой структурой. Хризокапсовые включают колониальные формы, неподвижные, пассивно плавающие или прикрепленные. Клетки этих организмов не имеют ни жгутиков, ни поверхностных выпячиваний. Они объединяются в единое целое общей слизью колоний, располагаясь обыкновенно в ее периферических слоях, однако могут находиться и в центральной части.
Класс Хризотриховые. К данному классу относятся золотистые водоросли, имеющие пластинчатую, нитчатую и разнонитчатую структуру. Все эти организмы являются многоклеточными, типично бентосными, прикрепленными. Таллом у них представлен разветвленными или простыми, одно- или многорядными нитями, дисковидными паренхиматозными пластиночками или кустиками. В общую слизь они не погружены. В этот класс объединяются пресноводные формы, реже морские и солоноватоводные. Хризотриховые являются самой высокоорганизованной группой организмов среди всех золотистых водорослей. Ее представители сходны по внешнему виду с улотриксовыми, относящимися к отделу Зеленых водорослей, а также с гетеротриксовыми, принадлежащими отделу Желто-зеленых водорослей. Отдельные представители хризотриховых напоминают некоторые наиболее просто устроенные бурые водоросли.
Класс Хризосферовые. К данному классу относятся золотистые водоросли, структура тела которых является коккоидной. Клетки этих организмов покрыты целлюлозной оболочкой. Жгуты и ризоподии у представителей данного класса полностью отсутствуют. Эти растения являются одноклеточными, неподвижными. Реже встречаются колониальные формы, которые представляют собой скопления клеток, слабо связанных между собой и не погруженных в общую слизь. Они не образуют пластинок или нитей при размножении.
Класс Хризофициевые. В этот класс объединяются золотистые водоросли, имеющие различные виды организации таллома. Именно его устройство является основанием, по которому в этом классе выделяются следующие порядки: ризохризидальные (имеющие ризоподиальную структуру), хризомонадальные (мопадные формы), хризокапсальные (пальмеллоидные формы), феотамниальные (нитчатые), а также хризосферальные (формы коккоидные). Предлагаем вам познакомиться с отдельными порядками данного класса. Хризомонадальные (иначе – хромулинальные) Это наиболее обширный порядок, объединяющий золотистые водоросли, имеющие монадную структуру, как колониальные, так и одноклеточные.
Систематика хризомонад основана на строении и количестве жгутиков. Особое значение имеет характер их клеточных покровов. Существуют формы одно- и двужгутиковые. Раньше считалось, что более примитивными, исходными являются именно первые. Однако электронный микроскоп помог ученым обнаружить, что у якобы одножгутиковых форм присутствует второй боковой жгутик небольшого размера. Исследователи предположили, что исходными могли быть двужгутиковые хризомонады, имеющие гетероморфные и гетероконтные жгутики, а одножгутиковые формы появились в результате последующей редукции короткого жгута. Что касается клеточных покровов представителей хризомонадальных, то они различны. Существуют голые формы, одетые исключительно плазмалеммой. Клетки других видов заключены в особые целлюлозные домики. Поверх плазмалеммы третьих находится покров, состоящий из окремнелых чешуек. С помощью деления клеток осуществляется процесс размножения хризомонад. У некоторых видов имеется и половой процесс. Следует отметить, что хризомонады – это в основном пресноводные организмы. Чаще всего они обитают в чистых водах. Хризомонады встречаются обычно в холодное время года, поздней осенью и ранней весной. Некоторые организмы обитают подо льдом в зимнее время. Однако, как выяснили ученые, для них не так важна температура воды. Она имеет только косвенное значение. Химизм воды является решающим фактором. На протяжении года он меняется: в холодное время года в воде в результате отсутствия другой растительности содержится больше азота и железа. Большинство хризомонад обитает в планктоне. У них есть особые приспособления для ведения планктонного образа жизни. Отдельные представители хризомонад окрашивают воду в бурый цвет, вызывая ее "цветение".
4 Разножгутиковые водоросли.
Для разножгутиковых (или Желто-зеленых) водорослей характерны следующие пигменты: хлорофиллы а, с, е (хлорофилл В отсутствует), Р-каротин, е-каротин, ксантофиллы: антераксантин, лютеин, зеаксантин, неоксантин, виолаксантин, вошериаксантин, гетероксантин, диадиноксантин, диатоксантин. Клетка содержит обычно по два пластинчатых хроматофора, которые лежат в цистерне эндоплазматической сети, находящейся в прямой связи с оболочкой ядра, в матриксе хлоропласта расположены трехтилакоидные ламеллы; почти всегда (ксантофициевые) имеется опоясывающая ламелла (отсутствует у эвстигматофициевых), внутренние пиреноиды ксантофициевых содержат трехтилакоидные ламеллы (рис 74). Глазок (стигма) является частью пластиды (ксантофициевые) или же гранулы стигмы располагаются вне пластиды (эвстигматофициевые).
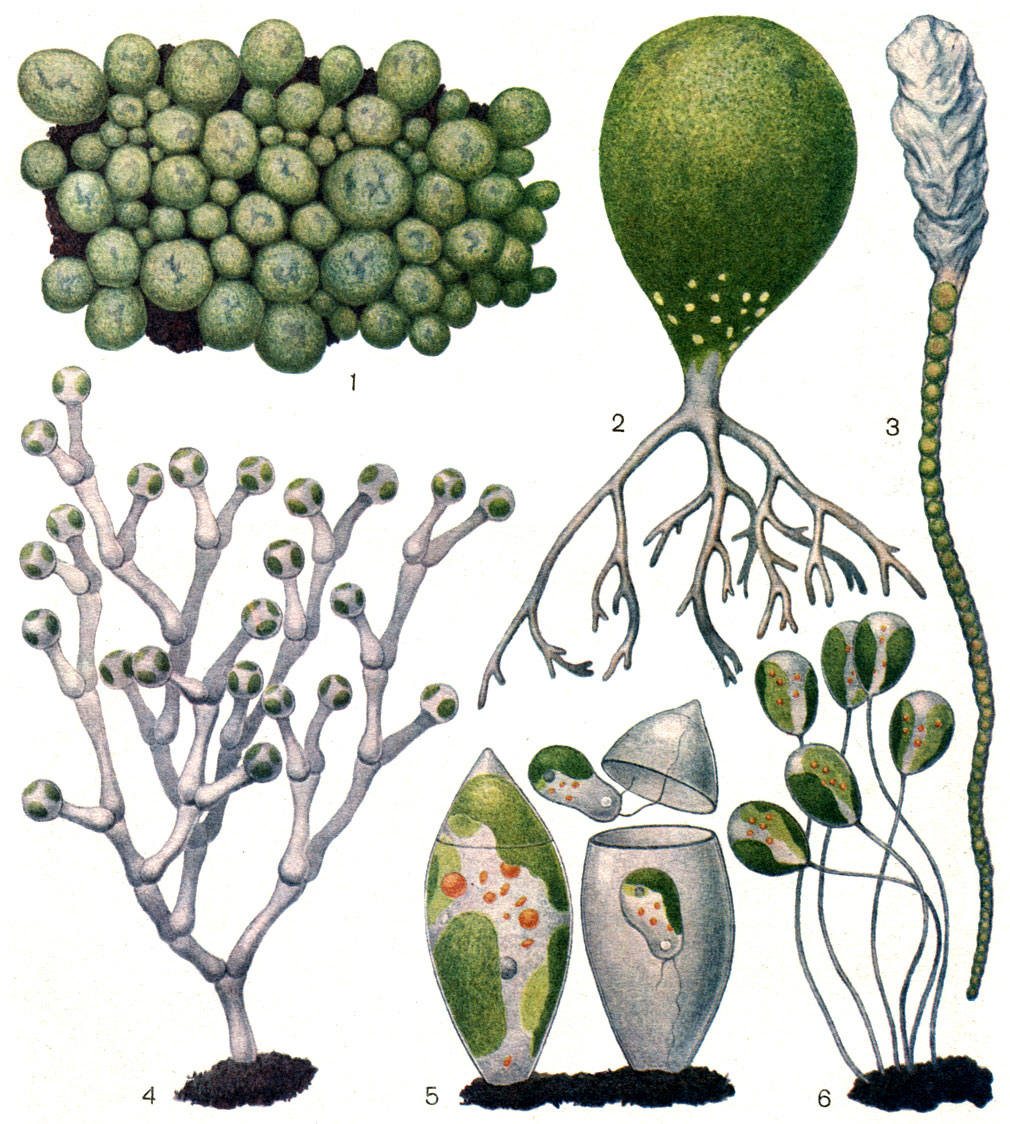
Рисунок 74 – Разножгутиковые водоросли
Запасные продукты: волютин, жир, часто хризоламинарин; крахмал не образуется. Более длинный жгутик несет мастигонемы, короткий - гладкий. Клеточные оболочки часто состоят из двух и более частей, цисты (статоспоры), как и у золотистых водорослей, эндоплазматические, их оболочки окременелые.
В большинстве современных систем различают два класса: ксантофициевые (Xanthophyceae) и эвстигматофициевые (Eustigmatophyceae).
Этот класс включает водоросли, стоящие на самых разных ступенях дифферешщровки таллома: монадные, ризоподиальные, лальмеллоидные, коккоидные, нитчатые, сифоновые. В соответствии с типами организации таллома различают порядки, параллельные порядкам золотистых, динофитовых, собственно зеленых водорослей. Порядок гетерохлоридальные (Heterochloridales) объединяет монадные формы, порядок ризохлоридальные (Rhizochioridales) включает ризоподиальные, порядок гетероглеальные (Helerogloeales) - пальмеллоидные, порядок мисхококкальные (Mischococcales) - коккоидные, порядок трибонемальные (Tribonematales) - нитчатые, порядок ботридиальные (Bo(rydiales) - сифоновые водоросли.
Порядок мисхококкальные – Mischococcales включает многочисленные одноклеточные и колониальные коккоидные формы. Клетки различных очертаний одеты клеточной стенкой, часто состоящей из двух частей. Размножение зооспорами или апланоспорами.
Род ботридиопсис (Botrydiopsis) широко распространен в пресноводных водоемах (прудах, канавах и пр.), представлен одиночными шаровидными клетками, одетыми клеточной оболочкой, под которой располагается постенная цитоплазма, содержащая многочисленные дисковидные хлоропласты, а в зрелых клетках - много клеточных ядер. Центр клетки занят крупной вакуолью с клеточным соком, пересеченной тонкими тяжами цитоплазмы. В цитоплазме рассеяны капли масла и глыбки хризоламинарина. Размножение зооспорами и автоспорами, образующимися в клетке в больших количествах (до 300). У В. arrhizaпри зооснорообразовании, как и у многих других желтозеленых водорослей (харациопсис, трибонема, ботридиум и др.), наряду с типичными зооспорами нередко наблюдаются синзооспоры.
Порядок трибонемальные – Tribonematales нитчатые формы. В качестве представителя может быть рассмотрен широко распространенный в пресных водах род трибонема (Tribonema). Таллом представлен неразветвленной, сложенной из одного ряда клеток нитью.
Цилиндрические, часто слегка бочонковидные клетки, обычно одноядерные, большей частью содержат несколько хлоропластов. В цитоплазме -жир, хризоламинарин. Клеточная стенка всегда состоит из двух половинок, краями находящих друг на друга в срединной плоскости клеток. Каждая половинка оболочки составлена из ряда налегающих друг на друга слоев.
Еще до деления ядра клетки (на стадии интерфазы) в экваториальной области под старой клеточной оболочкой закладывается новый цилиндрический кусок оболочки - промежуточное кольцо. На более поздней стадии деления материнской клетки, во время цитокинеза, в середине этого полого цилиндра образуется поперечная перегородка. Новая оболочка, имеющая в продольном оптическом сечении Н-образную форму, при разрастании раздвигает друг от друга обе более старые половинки материнской клеточной стенки, вставляясь между ними. Таким образом, оболочки соседних клеток соединяются друг с другом; соответственно каждая клетка окружена двумя половинками соседних Н-образных фигур. При освобождении репродуктивных клеток или под воздействием некоторых агентов (например, крепкой хромовой кислоты) оболочка клеток распадается на Н-образные участки. Фрагменты нитей всегда оканчиваются пустыми половинками Н-образных фигур, которые в профиль выглядят как два острия. Размножение нитей, непрерывно нарастающих за счет поперечных делений клеток, осуществляется вегетативно - фрагментацией и бесполым путем - посредством зооспор, амебоидных клеток, апланоспор, акинет.
Порядок ботридиальные – Botrydiales объединяет ксантофициевые водоросли, имеющие сифоновую организацию.
Представители рода ботридиум {Botrydium) живут на сырой земле. Таллом сифоновый, дифференцированный на надземную часть в виде сферического грушевидного пузыря диаметром 1-2 мм, и погруженную в субстрат ризоидальную часть, обычно разветвленную. Эта нормальная форма таллома в зависимости от внешних условий может заметно видоизменяться. Например, при культивировании ботридиума в погруженном в воду состоянии наблюдаются талломы в виде разветвленных нитей. Стенка многослойная, с различно ориентированными в каждом слое целлюлозными микрофибриллами. Цитоплазма сосредоточена в постенном слое и окружает непрерывную вакуоль с клеточным соком. Во взрослом талломе цитоплазма содержит многочисленные ядра, в надземной части - обильные дисковидные хлоропласты с пиреноидами и прочие органеллы. Размножение осуществляется одноядерными зооспорами с двумя гетероконтными и гетероморфными жгутиками, возникающими при погружении пузыря в воду (после дождя и т. п.). Наблюдались и синзооспоры. Оседая затем по краю лужи, на подсыхающей земле, зооспоры развиваются в новые растения. При более сухой погоде вместо зооспор образуются апланоспоры. По достижении известной внутренней зрелости ботридиум переходит в стадию покоя. Русский исследователь В. В. Миллер, подробно изучивший род ботридиум в культуре, описал различные способы образования покоящихся цист. В одних случаях все содержимое пузыря дает начало одной крупной цисте с толстой оболочкой. Такие цисты В. В. Миллер назвал «макроцистами» (В. walirothi, В. tuberosum, В. pachidermum). В других случаях содержимое надземной части делится с образованием многих многоядерных спороцист (В. walirothi). Наконец, все содержимое пузыря или часть его может переходить в ризоиды и там образовывать ризоцисты. Последние или располагаются по многу в ряд, например, у В. granulatum, или же содержимое переходит во вздутые концы ризоидов, в каждом из которых образуется циста, например у В. tuberosum. У одного и того же вида могут наблюдаться различные формы цист, которые при неодинаковых внешних условиях заменяют друг друга. Все формы цист для своего прорастания не нуждаются в периоде покоя, они могут прорастать и непосредственно после своего возникновения. Мелкие цисты (расположенные в ряд ризоцисты, спороцисты) или непосредственно прорастают в новые индивиды, или образуют зооспоры. Крупные цисты (макроцисты, образующиеся поодиночке ризоцисты В. tuberosum) прорастают обычно зооспорами или апланоспорами. Ни у одного из исследованных видов ботридиума В. В. Миллер не наблюдал полового процесса. Однако имеются данные других авторов о существовании у В. Granulatum полового процесса, изо- и гетерогамного у разных рас.
У рода вошерия (Vaucheria) (известны как пресноводные, так и морские и солоноводные виды) таллом в виде разветвленных сифоновых нитей, образующих ватообразные скопления в воде или обширные дерновинки темно-зеленого цвета на влажной земле. Рост нитей апикальный. В вегетативных нитях вошерии, начиная с верхушки, можно различить три зоны: апикальную, субапикальную и вакуолизированпую. Апикальный отрезок представляет собой активно растущую часть сифонового таллома: здесь находятся многочисленные пузырьки и митохондрии; хлоропласты и клеточные ядра отсутствуют. Везикулы содержат фиброзный материал, возможно, являющийся предшественником материала (целлюлозы) клеточной оболочки. В субапикальной зоне количество пузырьков уменьшается, появляются хлоропласты и клеточные ядра. Хлоропласты многочисленные, дисковидные, лишены пиреноидов. Выдающийся в виде почки пиреноид наблюдается только в проростках вошерии. С каждым клеточным ядром связана пара центриолей, которые во время профазы митоза мигрируют к противоположным полюсам удлиняющихся ядер. Оболочка ядра во время митоза остается интактной, внутри нее образуется внутриядерное веретено, центромеры отсутствуют; между расходящимися хромосомами ядерная оболочка перешнуровывается и окружает телефазные дочерние ядра. Нормальный митоз, происходящий внутри оболочки материнского ядра, у ряда видов вошерии наблюдал еще Л. И. Курсанов (1911). У всех исследованных видов Л.И. Курсанов отметил интересное распределение ядерных делений на протяжении нити: начавшись в одном каком-нибудь месте, митозы мало-помалу распространяются на соседние участки и т. д. Таким образом, на препарате, фиксирующем известный момент этого процесса, получается то, что всего лучше назвать волной делений, где отдельные, следующие друг за другом во времени фазы деления на протяжении нити расположены в правильной последовательности. В апикальной и субапикальной зонах еще отсутствует центральная вакуоль. Лишь в более старой - вакуолизированной - части нити появляется вакуоль с клеточным соком. Размножение бесполое, посредством синзооспор и апланоспор.
Класс эвстигматофициевые (Eustigmatophyceae) был выделен из ксантофициевых на основе строения монадных клеток (зооспор), в первую очередь их глазка. Глазок представляет собой крупное оранжево-красное тело у крайнего переднего конца зооспоры, не зависимое от единственной пластиды, и состоит из неправильной группы капель без ограничивающих мембран и без мембраны вокруг всего комплекса. Таким образом, в отличие от ксантофициевых стигма здесь расположена вне пластиды. Другой отличительный признак эветигматофициевых - то, что утолщение жгута, перекрывающее стигму, находится у проксимального конца, направленного вперед и несущего два ряда волосков жгутика. Гладкий, направленный назад жгутик, как правило, отсутствует. Кроме того, у эвстигматофициевых трехтилакоидные ламеллы часто образуют граноподобные стопки, периферическая опоясывающая ламелла отсутствует, пиреноид, обнаруженный только в вегетативных клетках (в зооспорах отсутствует), выступает от внутренней поверхности пластиды и не пересечен тилакоидами.
Все эвстигматофициевые - одноклеточные коккоидные формы (прежде относимые к порядку мисхококкальные из класса ксантофициевые).
Основная литература:
1 Еленевский А.Г., Соловьев М.П., Тихомиров В.Н. Ботаника: систематика высших, или наземных, растений. 2 изд. - М.: Academіa, 2001. - 429 с.
2 Нестерова С.Г. Лабораторный практикум по систематике растений. - Алматы: Қазақ ун-ті, 2011. - 220 с.
3 Родман А.С. Ботаника. – М.: Колос, 2001. - 328 с.
Дополнительная литература:
1 Абдрахманов О.А. Систематика низших растений. – Караганда: Изд-во КарГУ, 2009. - 188 с.
2 Билич Г.Л., Крыжановский В.А. Биология. Т. 2: Ботаника. - М.: Оникс 21 век, 2002. - 543 с.
3 Абдрахманов О.А. Практические работы по систематике низших растений. Ч. 2. Грибы и водоросли. – Караганда: Изд-во КарГУ, 2001. - 144 с.
4 Абдрахманов О.А. Лабораторный практикум по бактериям и водорослям. Учебное пособие. - Алматы: Казакадем образование, 2000. - 130 с.
5 Абдрахманов О.А. Практические работы по систематике низших растений. Ч. 2. Грибы и водоросли. – Караганда: Изд-во КарГУ, 2001. - 144 с.
Контрольные вопросы:
1 Какие черты организации характерны для водорослей? В чем их отличие от прокариот?
2 Дайте определение изогамии, гетерогамии и оогамии.
3 Какую роль играют водоросди на земле?
4 Какие основные систематические группы водорослей известны?
5 Какие представители характерны для золотистых водорослей?
6 Какие особенности строения положены в основу систематики разножгутиковых водорослей?
7 Что характерно для строения пиррофитовых водорослей?

