




Механизм ПЦР
ПЦР заключается в амплификации (умножении) в пробирке определенного участка ДНК в процессе повторяющихся температурных циклов (рис 2).
В основе ПЦР лежит обычная биохимическая реакция синтеза, катализируемая ферментом ДНК-полимеразой. Открытие ПЦР было обусловлено расшифровкой нуклеотидных последовательностей генома ряда микроорганизмов и обнаружением термоустойчивой Taq-ДНК-полимеразы. Данный фермент находится в живущих в горячих источниках бактериях Thermus aquaticus, обладает исключительной термостойкостью и сохраняет ферментативную активность в течение 40 минут при температуре 95° и имеет оптимум работы при 70–72°. ПЦР – это процесс, состоящий из многократно повторяющихся циклов и заканчивающийся получением большого количества копий специфического фрагмента ДНК.
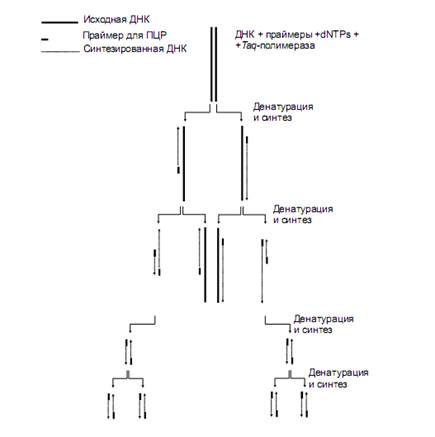
Рис.2. Диаграмма полимеразной цепной реакции (ПЦР). Полностью показаны только два первых цикла. Начиная с 3-го цикла, на диаграмме не отражена судьба исходной ДНК и продуктов достройки, синтезированных на ее цепях. Длинные фрагменты, синтезируемые на исходной цепи, накапливаются по формуле арифметической прогрессии. В противоположность этому - короткие дискретные фрагменты, ограниченные на концах праймерами, которые впервые появляются только в конце третьего цикла, накапливаются в геометрической прогрессии и очень скоро начинают доминировать среди продуктов амплификации
Ключевым компонентом реакции, помимо Taq-ДНК-полимеразы, являются короткие (20–30 нуклеотидов) искусственно синтезированные олигонуклеотиды – праймеры, комплементарные искомым участкам на левой и правой границах матричной ДНК. Праймеры выполняют ряд важных функций при проведении ПЦР:
1) праймеры являются точкой инициации синтеза новой цепи ДНК,
2) достраивание новой цепи ДНК протекает только между ними,
3) гибридная двуцепочечная система ДНК-праймер является местом присоединения ДНК-полимеразы к матрице.
ПЦР проводят в специальном программируемом термостате – амплификаторе, осуществляющем смену температурных циклов по заданной программе. Основным компонентом ДНК-амплификатора является элемент Пэльтье, способный осуществлять быстрые температурные переходы. Каждый цикл ПЦР состоит из трех этапов (денатурация, отжиг, элонгация).
Этапы ПЦР
1. Этап денатурации. Обычно, для денатурации (разрушения водородных связей и расхождения цепей ДНК) при 94°С достаточно 20-30 сек (отдельные авторы рекомендуют использовать 1-5 мин при ПЦР длинных фрагментов >5000 п.н.), но условия зависят от используемых пробирок и амплификатора. Если температура денатурации слишком низкая, не полностью денатурированная ДНК «схлопывается», что препятствует отжигу праймеров.
2. Этап отжига праймеров. При наличии в пробе искомого участка ДНК происходит присоединение (отжиг) праймеров (по одному на каждую цепь). Температура присоединения праймеров составляет в зависимости от их нуклеотидного состава 58–70°. В большинстве случаев температура отжига праймеров подбирается эмпирически. Выбор температуры отжига праймеров, возможно, один из самых ключевых факторов для обеспечения высокоспецифичной ПЦР.
Если температура окажется завышенной, отжиг не будет происходить, если будет занижена - резко увеличится неспецифический отжиг. Если праймер имеет 3'-концевую область, имеющий гомологию к какому-то другому участку на матрице, праймер, вне зависимости от того, специфически он отжигается или нет, будет читаться с этой 3'- концевой области. При низкой температуре возможен и такой вариант, когда праймер гибридизуется без участия 3'-конца и в таком случае чтение с праймера проходить не будет. Праймер-димеры образуются, если праймеры имеют одно или более комплементарных оснований на 3'-конце, так, что становится возможным комплементация между двумя 3'-концами двух праймеров.
3.Этап элонгации (синтеза). При подъеме температуры до 72°С активируется Taq-ДНК-полимераза и начинается этап синтеза, заключающийся в присоединении дезоксинуклеотид-трифосфатов и достраивании фрагмента ДНК. Чтение с праймеров нормально проходит при 72°С - температурном оптимуме для Taq-полимеразы. Обычно, 20 сек вполне достаточно для эффективного синтеза 500 п.н. и 40 сек - для фрагмента 1.2 т.п.н. Однако, время реакции может меняться в зависимости от используемого фермента и величины получаемого фрагмента (например, Taq-полимераза присоединяет 60-200 оснований в секунду при 72 °С).
Перед началом цикличного процесса проводят предварительную денатурацию. Очень важно, чтобы ДНК, которая выступает в качестве матрицы, была полностью денатурирована. Обычно нагрева до 94°С или 95°С в течении 4 минут достаточно для полного разделения цепей ДНК.
Число циклов. В условиях оптимальной реакции для выявления целевой последовательности ДНК, достаточно не более 40 циклов амплификации, чтобы без труда визуализировать в геле с этидиумом бромидом полученный в ходе ПЦР продукт. В большинстве же случаев достаточно всего 25-30 циклов. Следует помнить, что с увеличением числа циклов идет накопление неспецифического продукта. В большинстве случаев, минимальным числом копий, дающих стабильно воспроизводимый результат в ходе ПЦР служит 100 - 1000 копий ДНК- матрицы.
Конечная элонгация. Обычно, после последнего цикла, реакционную смесь инкубируют дополнительно в течении 5-15 мин при 72 °С для достройки частично достроенных продуктов.
Образовавшиеся в первом цикле амплификации новые цепи ДНК служат матрицами для второго цикла амплификации, в котором происходит образование искомого специфического фрагмента ДНК (ампликона) (Рис.3.). С каждым последующим циклом количество копий участка ДНК удваивается. Таким образом, происходит накопление ампликонов по формуле:
х = 2n, где n – число циклов.
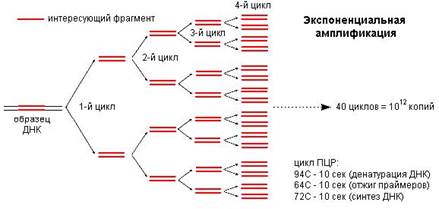
Рис.3 Экспоненциальный рост ампликонов в ходе ПЦР. (Иллюстрированный материал с сайта www.dna-technology.ru).
В результате 30–40 циклов в растворе накапливается около 100 миллионов молекул ампликона. Этого количества достаточно для визуальной детекции этого фрагмента методом электрофореза в агарозном геле, содержащем бромистый этидий. Реальное значение эффективности отдельных циклов амплификации составляет по некоторым данным 78-97%. В случае присутствия в пробе ингибиторов реакции это значение может быть намного меньше, поэтому фактическое количество специфических продуктов амплификации лучше описывает формула:
А = М × n(1+Е), где
А – количество специфических (ограниченных праймерами) продуктов реакции амплификации; М – начальное количество ДНК-мишеней; n – число циклов амплификации; Е – значение эффективности реакции.
Следует заметить, что в процессе амплификации на исходной цепи синтезируются и длинные фрагменты, однако их накопление происходит лишь в арифметической прогрессии по формуле К = М × n, где К – количество длинных продуктов амплификации.
Таким образом, специфические фрагменты, ограниченные на концах праймерами, впервые появляются в конце второго цикла, накапливаются в геометрической прогрессии и очень скоро начинают доминировать среди продуктов амплификации.
«Эффект плато». Следует заметить, что процесс накопления специфических продуктов амплификации по геометрической прогрессии идет лишь ограниченное время, а затем его эффективность критически падает. Это связано с так называемым «эффектом плато». Термин «эффект плато» используют для описания процесса накопления продуктов ПЦР на последних циклах амплификации, когда количество ампликонов достигает 0,3 - 1 пмолей.
В зависимости от условий и количества циклов реакции амплификации, на момент достижения «эффекта плато» влияют: утилизация субстратов (дНТФ и праймеров); стабильность реагентов (дНТФ и фермента); количество ингибиторов, включая пирофосфаты и ДНК-дуплексы; неспецифические продукты или праймер-димеры, конкурирующие за праймеры, дНТФ и полимераза; концентрация специфического продукта и неполная денатурация при высокой концентрации продуктов амплификации. Чем меньше начальная концентрация ДНК-мишени, тем выше риск выхода реакции на плато. Этот момент может наступить до того, как количество специфических продуктов амплификации будет достаточно, чтобы их можно было проанализировать.
Существуют различные модификации ПЦР, которые используются в зависимости от конкретных целей проведения реакции или от характера последующего молекулярного анализа амплификатов. Так, для трудно амплифицируемых участков ДНК (содержащих различные повторяющиеся последовательности или необычные структурные элементы), а также в тех случаях, когда матричная ДНК присутствует в следовых количествах, ПЦР проводят в два раунда, используя в качестве матричной ДНК на втором этапе амплификации, или как еще говорят при доамплификации, продукты ПЦР, синтезированные в первом раунде. Часто в этих случаях для повышения специфичности праймирования используют систему, так называемых вмонтированных (nested) праймеров, то есть при доамплификации в качестве праймеров выбирают последовательности, локализованные внутри амплифицируемого в первом раунде участка ДНК.
В ряде случаев удобно проводить мультиплексную ПЦР, то есть одновременную амплификацию нескольких участков матричной ДНК. Можно получать меченые продукты ПЦР, добавляя в реакционную смесь меченые трифосфаты. Особого внимания заслуживает возможность проведения ПЦР с молекулами кДНК.
На основе этой реакции разработаны методы анализа экспрессии генов и получения больших количеств кДНК. Уместно заметить, что реакцию амплификации можно проводить не только в растворах, но и непосредственно на хромосомных препаратах, при этом в случае использования меченых нуклеотидов продукты амплификации будут гибридизоваться и выявлять комплементарные им участки ДНК на хромосомах (метод PRINS - polymerase reaction in situ). До настоящего времени доступными амплификации были участки ДНК, не превышающие по длине 5 т.п.н. В последнее время, благодаря внесению ряда кардинальных усовершенствований (особый подбор праймеров, использование сразу двух различных ДНК полимераз, трицинового буфера, специального температурного режима полимеразных циклов), возможно проведение амплификации фрагментов ДНК, достигающих 35 т.п.н. В частности, таким образом удалось амплифицировать плазмиду со вставкой 8.5 т.п.н., равной целому геному вируса HIV 1. И это еще не предел. В дальнейшем мы неоднократно будем возвращаться к описанию различных модификаций ПЦР, так как этот метод по праву стал один из основных в молекулярной диагностике наследственных болезней.
Возможность очень точного и специфичного выбора участка ДНК для амплификации, небольшие размеры синтезируемых молекул, их огромное количество облегчают молекулярный анализ продуктов ПЦР. Как правило, амплифицированную ДНК можно непосредственно наблюдать в проходящем ултрофиолете в виде красной полосы после электрофоретического концентрирования и обычного окрашивания геля этидиумом бромидом (рис.4).
Детекцию продуктов ПЦР-реакции проводят с помощью метода гель-электрофореза ДНК (См. гл.3.)
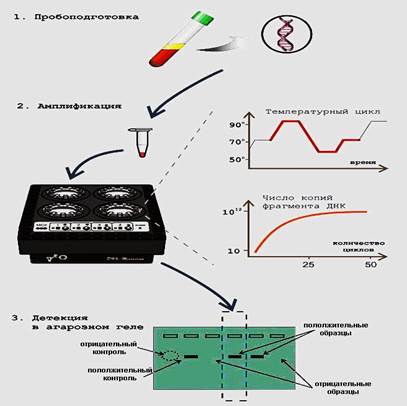
Рис. 4. Обобщенная схема ПЦР с последующей детекцией в агарозном геле (иллюстрированный материал с сайта www.dna-technology.ru).

