




Эффекторный нейрон. Для соматической нервной системы это мотонейрон. Его назначение - вместе с другими нейронами ЦНС
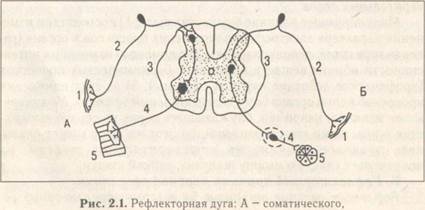 Б - вегетативного рефлексов
Б - вегетативного рефлексов
|
Переработать информацию, сформировать ответ в виде нервных импульсов, посылаемых к 5 звену рефлекторной дуги.
5. Эффектор (рабочий орган). В простейшем случае рефлекторная дуга может быть двухнейронной - без вставочного нейрона.
В. Классификация рефлексов проводится по нескольким критериям. В частности, по срокам появления рефлексов в онтогенезе их делят на две группы: врожденные (безусловные) и приобретенные (условные). Безусловные рефлексы делят также на несколько групп. По биологическому значению рефлексы делят на пищедобывательные, половые, защитные (оборонительные) и др. В зависимости от отдела нервной системы выделяют соматические и вегетативные рефлексы.
Г. Нервизм - это концепция, признающая ведущую роль нервной системы в регуляции функций всех органов и тканей организма. Выдающийся вклад в развитие концепции физиологического нервизма сделал И. П. Павлов (1849-1936). Он обосновал представление о трофическом влиянии нервной системы на органы и ткани, сформулировал принципы рефлекторной теории, доказал важную роль нервной системы в регуляции секреции желез желу- дочно-кишечного тракта, открыл условные рефлексы и с их помощью разработал основы учения о высшей нервной деятельности.
Д. Нервный центр - это совокупность нейронов, расположенных на различных уровнях ЦНС, достаточных для приспособительной регуляции функции органа согласно потребностям организма. Например, нейроны дыхательного центра располагаются и в спинном мозге, и в продолговатом мозге, и в мосту. Однако среди нескольких групп клеток, расположенных на различных уровнях ЦНС, обычно имеется главная часть центра. Главная часть дыхательного центра находится в продолговатом мозге и включает инспираторные и экспираторные нейроны.
Рассмотрим более подробно отдельные элементы рефлекторной дуги.
Сенсорные рецепторы как первое звено рефлекторной дуги
Различают эффекторные и сенсорные рецепторы.
Эффекторные рецепторы (лат. recipere - получать) представляют собой белковые структуры клеточных мембран, а также цитоплазмы и ядра, активируются химическими соединениями (медиаторами, гормонами), что запускает ответные реакции клетки.
Сенсорные рецепторы воспринимают раздражители внутренней и внешней среды организма с помощью трансформации энергии раздражения в нервный импульс. Их раздражителями являются изменение температуры, прикосновение, давление, изменение рН, осмотического давления и т.д. Основное физиологическое значение сенсорных рецепторов состоит в обеспечении поступления в ЦНС информации о состоянии внешней и внутренней среды, что обеспечивает регуляцию функций внутренних органов и организацию взаимодействия организма и окружающей среды, поддержание тонуса ЦНС.
А. Классификация сенсорных рецепторов.
1. По структурно-функциональной организации различают первичные и вторичные рецепторы. Первичные рецепторы представляют собой чувствительные окончания дендрита афферентного нейрона (рис. 2.2). Его тело локализуется в спинномозговых ганглиях или в ганглиях черепных нервов. Афферентные нейроны локализуются также в вегетативных ганглиях. К первичным рецепторам относятся тактильные, болевые, температурные, проприоре- цепторы, обонятельные рецепторы, механо- и хеморецепторы внутренних органов. Вторичные рецепторы имеют специальную клетку, синаптически связанную с окончанием дендрита афферентного нейрона (см. рис. 2.2). К вторичным рецепторам относятся вестибулярные, слуховые, фоторецепторы и вкусовые рецепторы.
2. С психофизиологической точки зрения рецепторы подразделяются в соответствии с органами чувств и формируемыми ощущениями на зрительные, слуховые, вкусовые, обонятельные и тактильные.
3. В зависимости от вида воспринимаемого раздражителя выделяют пять типов рецепторов. Механорецепторы расположены в коже, внутренних органах, сосудах, слуховой и вестибулярной системах, опорно-двигательном аппарате. Хеморецепторы локализуются в слизистой оболочке носа, языка, каротидном и аортальном тельцах, продолговатом мозге и гипоталамусе. Терморецепторы (тепловые и холодовые) расположены в коже, сосудах, внутренних органах, гипоталамусе, продолговатом, спинном и среднем мозге, фоторецепторы - в сетчатке глаза. Болевые рецепторы (ноцицепторы) — их раздражителями являются механические, термические и химические (гистамин, брадикинин, К+, Н+ и другие вещества) факторы - локализуются в коже, мышцах, внутренних органах, сосудах, дентине.
4. По расположению в организме различают экстеро- и инте- рорецепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек и органов чувств: зрительные, слуховые, вкусовые, обонятельные, тактильные, кожные, болевые и температурные. К интерорецепторам относят рецепторы внутренних органов, рецепторы опорно-двигательного аппарата (проприорецепторы) и вестибулорецепторы. Имеются и другие классификации.
Б. Механизм возбуждения рецепторов.
Возникновение рецепторного потенциала (РП). При действии на рецептор адекватного раздражителя (к которому он эво- люционно приспособлен), способного вызывать конформационные изменения воспринимающих структур (активацию рецепторного белка), формируется рецепторный потенциал. В рецепторах (кроме фоторецепторов) энергия раздражителя, после ее преобразования и усиления, приводит к открыванию ионных каналов и перемещению ионов, среди которых основную роль играет движение Na+ в клетку. Это приводит к деполяризации мембраны рецептора. Полагают, что в механорецепторах растяжение мембраны ведет к расширению каналов. Рецепторный потенциал является локальным, он может распространяться только электротонически на небольшие расстояния - до 3 мм. Возникновение потенциала действия (ПД) в первичных и вторичных рецепторах происходит по-разному.
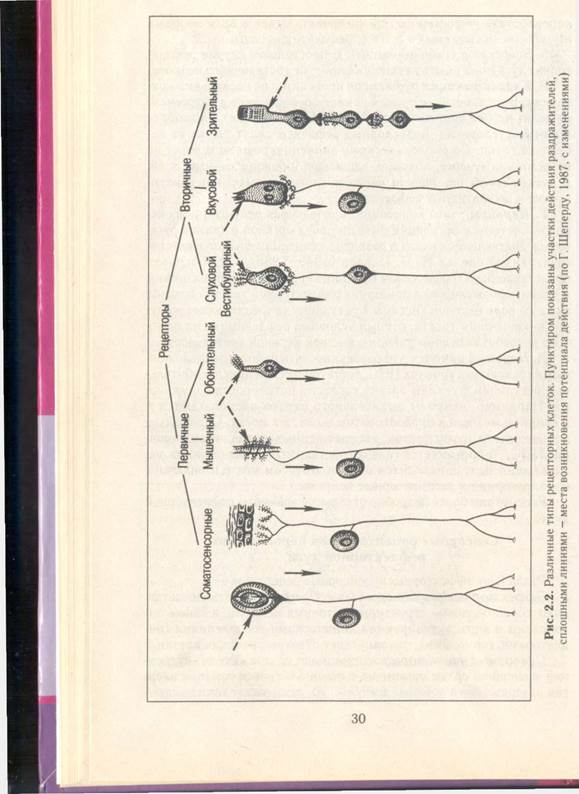
|
Возникновение ПД. В первичном рецепторе рецепторная зона является частью афферентного нейрона - окончанием его дендрита. Она прилежит к рецептору. Возникший РП, распространяясь электротонически, вызывает деполяризацию нервного окончания и возникновение ПД. В миелиновых волокнах ПД возникает в ближайших перехватах Ранвье, в безмиелиновых - в ближайших участках, имеющих достаточную концентрацию потенци- алзависимых натриевых и калиевых каналов, а при коротких дендритах (например, в обонятельных клетках) - в аксонном холмике. При достижении деполяризации мембраны критического уровня происходит генерация ПД. Во вторичных рецепторах РП возникает в рецепторной клетке, синаптически связанной с окончанием дендрита афферентного нейрона. РП обеспечивает выделение рецепторной клеткой медиатора в синаптическую щель. Под влиянием медиатора на постсинаптической мембране возникает генераторный потенциал, обеспечивающий возникновение ПД в нервном окончании вблизи постсинаптической мембраны. Генераторный потенциал, как и рецепторный, является локальным потенциалом.
В. Свойства рецепторов.
1. Высокая возбудимость рецепторов. Например, для возбуждения фоторецептора сетчатки достаточно одного кванта света, для обонятельного рецептора - одной молекулы пахучего вещества.
Адаптация рецепторов - уменьшение их возбудимости при длительном действии раздражителя (только темновая адаптация фоторецепторов - это повышение их возбудимости). Адаптация рецепторов выражается в снижении амплитуды РП и, как следствие, в уменьшении частоты импульсации в афферентном волокне.
3. Спонтанная активность рецепторов, т.е. способность возбуждаться без действия раздражителя, присуща проприорецепто- рам, фоно-, фото-, вестибуло-, термо-, хеморецепторам. Эта способность связана со спонтанным колебанием проницаемости клеточной мембраны, перемещением ионов и периодической деполяризацией рецептора, которая, достигая критического уровня, приводит к генерации ПД в афферентном нейроне. Возбудимость рецепторов, обладающих фоновой активностью, выше, даже слабый раздражитель способен значительно повысить частоту импульсации в них. Фоновая активность рецепторов участвует в поддержании тонуса ЦНС.
Нервное волокно как элемент рефлекторной дуги
А. Классификация нервных волокон. Имеется два типа нервных волокон: миелиновые и безмиелиновые. Оболочку безмие- линовых волокон образуют шванновские клетки (леммоциты). Оболочку миелиновых волокон в периферической нервной системе формируют шванновские клетки, а в ЦНС - олигодендроци- ты. Миелиновая оболочка через равные промежутки (0,5-2,0 мм) прерывается, образуя свободные от миелина участки - узловые перехваты Ранвье, протяженность которых в волокнах периферической нервной системы составляет 0,25-1,0 мкм, в волокнах ЦНС их длина достигает 14 мкм. Миелиновая оболочка нервных волокон выполняет изолирующую функцию, обеспечивает более экономное и быстрое проведение возбуждения.
Классификация нервных волокон осуществляется согласно структурно-функциональным свойствам. В зависимости от толщины нервных волокон, наличия или отсутствия у них миелиновой оболочки все нервные волокна делят на три основных типа: А, В и С (Дж. Эрлангер и X. Гассер). Волокна типа А - это афферентные и эфферентные волокна соматической нервной системы, скорость проведения возбуждения 20-120 м/с; волокна типа В - преганг- лионарные волокна вегетативной нервной системы, скорость проведения возбуждения 5-10 м/с; волокна типа С - это в основном постганглионарные волокна вегетативной нервной системы, скорость проведения возбуждения 0,5-2 м/с.

