




Олар екі топқа бөлінеді: I топ – азотқұрамды протеин емес компоненттер. Қанның протеин емес азоты заттарының құрамына жай және күрделі протеиндердің аралық және соңғы алмасуларының өнімдері кіреді. Бұрын протеин емес азотқұрамдыларды «қалдық азот» деп атаған (олар протеинді тұндырғаннан кейін қалады). Олар мочевина азотынан (50%), аминоқышқылдар азотынан (25%), кіші молекулалы пептидтерден, креатиннен, креатининнен, билирубиннен, индиканнан, кейбір басқа да азотқұрамды заттардан тұрады.
Кейбір бүйрек аурулары кезінде, сонымен бірге, протеиндердің массивті ыдырауымен жүретін патологияларда (мысалы, ауқымды ауыр күюлерде), қанда протеинсіз азоттың мөлшері жоғарлайды, немесе, азотемия байқалады. Бірақ, жиі түрде протеинсіз азоттың жалпы мөлшері емес, протеинсіз азоттың жекеленген компоненттері арасындағы қатынас өзгереді. Сондықтан, қазіргі кезде, плазмадағы азоттың жеке компоненттерін анықтайды.
«Қалдық азот» түсінігіне кіші молекулалы пептидтерді де қосады. Кіші молекулалы пептидтерде жоғары биологиялық активтілік көрсететіндері де бар (мысалы, пептидтік табиғаттағы гормондар).
II топ – азотсыз органикалық заттар. Оларға көмірсулар, липидтер және олардың метаболизмінің өнімдері (глюкоза, пируват, лактат, кетондық денелер, май қышқылдары, холестерин және оның эфирлері) жатқызылады.
Плазманың бейорганикалық компоненттері
Қан құрамында келесі электролиттер болады: бейорганикалық катиондар - Na+, K+, Ca++, Mg++; бейорганикалық аниондар - Cl-, HCO3-, PO43-, HPO42-, H2PO4-; органикалық аниондар.
Электролиттердің қызметі: қанның осмостық қысымының болуын қамтамассыз етеді; қанның буферлік жүйесінің құрамына кіреді; қанның электролиттері ұлпа сұйықтықтары мен жасушалардың иондық құрамының тұрақтылығын ұстау үшін қажет.
Плазманың қышқылдық-сілтілік балансы
Плазмадағы және жасуша аралық кеңістіктегі ион Н+ концентрациясы 40 нМ-ге жуық болады. Бұл рН-тың 7,40 мәніне сәйкес келеді. Ағзаның ішкі ортасындағы рН мәні тұрақты болуы керек, себебі, протондар концентрациясының айқын өзгерістері өлімге әкелуі мүмкін. (сур. 2)
рН мәнінің тұрақтылығы плазманың буферлік жүйелері арқылы ұсталынады (схема В). Оларды қышқылдық-сілтілік баланстың қысқа мерзімді бұзылысы компенсациялайды. рН-тың ұзақмерзімді тепе-теңдігі протондар өндірілуі мен жойылуы арқылы ұсталынады. Буферлік жүйенің бұзылысы кезінде және қышқылдық-сілтілік баланс сақталмаған кезде, мысалы, бүйрек ауруларының нәтижесінде немесе тыныс алу кезеңінің бұзылысында (гипо-, немесе гипервентиляция нәтижесінде) плазма рН-ы қалыпты шектен асып кетеді. рН мәнінің 7,40-тан 0,03 бірлікке ауытқуының өзі ағзаны өлімге әкелуі мүмкін, төмен болса – ацидоз, ал жоғары болса алкалоз деп аталады.
Протондардың шығу көздері. Протондардың шығу көздері екеу: - тағамдағы бос қышқылдар (мысалы, лимон, аскорбин және фосфор қышқылдары, рН сілтілік болған кезде, ішек трактісінде протондарды бөледі) және тағам құрамында түсетін протеиндердің күкірт құрамды аминоқышқылдары. Протондар балансын қамтамассыз етуге протеиндер ыдыраған кезде түзілетін аминоқышқылдар метионин және цистеин үлес қосады. Бауырда бұл аминоқышқылдардың күкірті күкірт қышқылына дейін тотығады. Ол сульфат ионға және протонға диссоциацияланады.
Бұлшықет пен эритроциттердің анаэробтық гликолизі кезінде глюкоза сүт қышқылына айналады, оның диссоциациясы лактат пен протондардың түзілуіне әкеледі.
Кетондық денелер: -ацетоацетат және β-гидроксибутират – түзілуі (бауырда) де протондардың пайда болуына әкеледі. Кетондық денелердің артық мөлшері плазманың буферлік жүйесінің артық жүктелуіне әкеліп рН-тың (метаболиттік ацидоз пайда болады, сүт қышқылынан – лактацидоз, кетондық денелерден – кетоацидоз) дамиды. Қалыпты жағдайда бұл қышқылдар СО2 мен Н2О-ға дайін метаболиттенеді және протондық балансқа ықпал етпейді.
Протондардың жойылуы. Бүйректерде протондар Nа+ ионымен активті алмасып несеп құрамына өтеді. Ары қарай, несепте, протондар NН3-пен және фосфатпен әрекеттесіп буферленеді.
Плазманың буферлік жүйесі
Плазманың біршама маңызды буферлік жүйесі бикарбонаттық буфер болып табылады. Ол әлсіз көмір қышқылынан (рК1= 6,1) және оның қышқылдық анионынан – бикарбонаттан тұрады (сур.2.). Көмір қышқылы Н2СО3 өзінің ангидриді СО2 –мен тепе теңдікте болады. Екі форма арасындағы тепе теңдіктің тұрақтануы карбонатдегидрогеназа ("карбоангидраза") ферментімен сақталынады. Плазманың тұрақты рН мәнінде НСО3- және СО2 концентрациясы 20/1 қатынасындай болады. Қанда еріген СО2 өкпе альвеолаларында газдық фазадағы СО2-мен тепе теңдік жағдайда алмасады. Сондықтан НСО3-/СО2 –жүйесі тиімді ашық буферлік жүйе болып табылады. Үдемелі немесе баяу тыныс алу СО2 концентрациясын өзгертеді, бұл плазма рН-ы өзгеруіне әкеледі (тыныс алу ацидозы немесе сәйкес түрдегі алкалозға). Сондықтан, өкпелер жылдам және әрекетті түрде плазманың рН-на протондарды бөліп алу жүйесінің қатысынсыз да ықпал ете алады.
Қан плазмасының протеиндері, негізінен, эритроциттердің гемоглобині де протондарды қосып алуға қабілетті, сондықтан оларда рН тұрақтылығын ұстауға қатысады. Қанның буферлік жүйесінің қызметіне фосфаттар да белгілі бір үлес қосады.
ҚАННЫҢ НЕГІЗГІ ПРОТЕОЛИТТІК ЖҮЙЕЛЕРІ
Қан плазмасының құрамында бірнеше протеолиттік жүйе болады. Бұл жүйелердің құрамында – ағзаның қорғаныстық және реттеушілік реакцияларына қатысатын протеазалар кіреді. Ұлпалықтар мен салыстырғанда, плазмалық протеиназалар кеңістіктік таралып бөлінбеген. Сондықтан, олар бір бірімен еркін әсерлесе алады.
Плазмалық протеиназалар белсенуі «гетерогендік катализ» деген жалпы атауға біріктірілген үдерістер топтарына жатқызылады және бөтен беткейлермен байланысқан кезде әсерлі өтеді.
Қанның негізгі протеолиттік жүйелеріне жатқызылады:
1. қанның ұю жүйесі және фибринолиз
2. комплемент жүйесі – ағзаның иммундық қорғанысының құрам бөліктерінің бірі
3. кининдік жүйе
4. ренин-ангиотензиндік жүйе
Бұл жүйелер әртүрлі қызметтерді қамтамассыз етеді, бірақ олардың жұмысында протеолиттік жүйе қызметінің жалпы принципі сақталады. Олар:
1. Олар көпкомпонентті мультиферменттік жүйелер, олардағы бастапқы реакциялардың өнімдері келесі реакциялар үшін фермент болып табылады.
2. Бұл жүйелердің көптеген компоненттері протеолиттік ферменттер болып табылады. Олар проферменттер түрінде қанда айналады және тек белгілі бір жағдайда ғана активтенеді.
3. Бұл жүйелер бірінші ретті әлсіз сигналдарды күшейту қасиетінде болады. Олар каскадтар принципі бойынша жұмыс істейді немесе олардың жұмысы ферменттердің активті формалары мөлшерінің жылдам жоғарлауына әкеледі.
4. Жүйелер өзін-өзі реттеуді оң және кері теріс байланыстар принципі бойынша атқарады.
ҚАННЫҢ ҰЮ ЖҮЙЕСІ ЖӘНЕ ФИБРИНОЛИЗ
Бұл келесі маңызды қызметтерді атқаратын бірегей жүйе: 1) Тамырларда қанды сұйық күйде ұстау; 2) Гемостазды ұстау (көп қан кетуден қорғау).
Гемостаз - күрделі, ферменттік үдеріс, оның нәтижесінде қан ұйымасы түзіледі. Гемостазға 3 өзара байланысты механизм кіреді: 1. Тамырлық-тромбоцитарлық механизм; 2. Коагуляциялық механизм; 3. Тромб ретракциясы.
Тамырлық-тромбоцитарлық механизм тамыр зақымдалған соң алғашқы минутта активтенеді. Тамыр зақымдалған аймақта тромбоциттер жинақталады және жарақаттың жиектеріндегі эндотелийлерге жабысады. Тромбоциттер адгезиясының екі механизмі кездеседі: 1) Зақымдалған эндотелий мембранасының беткейі оң зарядталған қалыпқа ауыса бастайды. Сондықтан, оған беткейлері теріс зарядталған тромбоциттер жабысады. 2) Тамырдардың зақымдалуы бос Виллебранд факторының түзілуіне әкеледі (қалыпты жағдайда ол VIII фактормен ассоцияцияланған болады). Виллебранд факторы субэндотелиальді құрылымдар мен тромбоцит бетіндегі протеиндер арасында көпірше байланыстарды түзеді.
Тромбоциттердің адгезиясы олардың активтенуіне әкеледі. Активтелген тромбоциттер серотонинді, катехоламиндерді, АДФ-ті секреттейді. Серотонин тамыр тарылтырушы әсер береді. АДФ әсерінен тромбоциттер үймеленіп жиналады және борпылдақ тромбоциттік тығынды түзеді. Ол арқылы қан плазмасы өте алады. Осы мезгілде қан плазмасында түзілетін тромбин тромбоциттердің рецепторларына әсер етеді және олардың ыдырап тығыз массаға айналуын қамтамассыз етеді. Бұл түзілген тромбициттік тығын қан плазмасын өткізбейді.
Коагуляциялық механизм тамыр зақымдалған соң бірнеше минуттан кейін активтенеді. Коагуляция үдерісі - бұл сұйық қан плазмасының фибрин протеині негізінде тығыз гельге айналуы болып табылады. Түзілген гельді түйіршіктер тромбоциттік тығынды нығыздап күшейтеді.
Тромб ретракциясы – ұйыманың фибрин талшықтары және тромбоциттік тромбостенин есебінен сығылуы. Ретракция есебінен ұйыманың тығыздалуы және жарақат жиектерінің тартылуы өтеді.
Қанның ұю жүйесі немесе гемокоагуляция – бұл көпкомпонентті жүйе, оның құрамына протеиндер, фосфолипидтер, жасуша мембранасының сынықтары және кальций ионы кіреді.
Қанның ұю жүйесінің компоненттерін «факторлар» деп атау қабылданған. Факторлар ұлпалық, плазмалық, тромбоциттік болып бөлінеді. Ұлпалық және плазмалық факторлар рим цифрасымен, ал тромбоциттіктер – араб цифрасымен белгіленеді. Егер фактор активті болса, онда цифраның жанына "а" әріпі қойылады. Мысалы, активсіз он екінші фактордың активтіге айналуы келесі түрде белгіленеді: фXII ----> фXIIa
Қанның ұю жүйесінің көптеген протеиндері ферменттік активтілікті көрсетеді. Қанның ұюының барлық факторлары, фXIII-ден басқасы, сериндік протеиназа болып табылады, олар шектеулі протеолиз реакциясын катализдейді.
Қанның ұю реакциясында барлық протеин-ферменттер алғашқыда субстрат ролінде болады, сонан кейін фермент ролінде. Қанның ұюына қатысатын протеинтардың арасындаферменттік активтілік көрсетпейтіндері де болады. Олар ферменттік реакцияны спецификалық түрде күшейтеді. Олар параферменттер деп аталады. Параферменттер - фV және фVIII.
Қанның ұюының көптеген факторлары активсіз профермент күйінде синтезделеді. Айналымдағы қанда протеолиттік жүйенің проферменттері: VII фактор (проконвертин), IX фактор (Кристмас факторы), Х факторы (Стюарт факторы) және П фактор (протромбин).
Проферменттер активтеледі және олардың әсері қанның ұюының тікелей реакциясының жүруіне немесе қан ұйымасының негізі болатын фибриногеннің фибринге айналуына бағытталады.
Фибриннің түзілуі үш этапта өтеді. Алғашқыда мембранамен байланысқан протеазалар комплексі мен протеиндік факторлар түзіледі, ол протромбиназалық қасиетті көрсетеді.
Сонан кейін, протромбиназалық комплекс протеин-алғызат протромбинді тромбинге айналдырады.
Тромбин, өз кезегінде, фибриногенге фермент тәрізді әсер етеді де оны фибрин-мономерге айналдырады.
Мономердің фибрин-полимерге полимерленуі және фибрин-полимерде көлденең байланыстардың тігілуі XIII фактордың әсерінен жүріп тромбтың фибриндік бөлігі қалыптасады.
Протромбиназалық комплекстің түзілуі – фибриногенездің өте күрделі және ұзақ этапы болып табылады.
Ол ұлпа зақымдалуына жауап ретінде – ұюдың сыртқы каскады (механизмі) немесе плазма компоненттерінің полианиондармен жанасуына – ұюдың ішкі каскады (механизмі) ретінде де атқарылады.
Сыртқы (немесе тамырдан тыс) механизм сыртқы (ұлпалық) факторлардың қатысуымен босатылады. Ішкі механизм (немесе тамырішілік) – шығу көзі қанның өзі, плазма, қанның формалық элементтері, ферменттері болатын факторлардың қатысуымен босатылады.
Сыртқы каскадтың активтелуі кезінде мембраналық арқалық қызметін тамыр қабырғалары атқарады, ал ішкі каскад реакцияларында – қанның формалық элементері мембранасының фосфолипидтері атқарады.
Екі каскад та негізгі бір этапта - Х фактордың активтелуінде бірігеді.(Сур.2).
ҰЮДЫҢ СЫРТҚЫ МЕХАНИЗМІНІҢ БАСТАПҚЫ СТАДИЯЛАРЫ
Ұюдыңсыртқы механизмі босатылуы (басталуы) үшін бастапқы сигнал: - қанмен жанасушы ұлпаның немесе тамыр эндотелийі болуы қажет. Бұл жағдайда жасуша мембранасы ыдырайды. Жасушалардан ұлпалық тромбопластин (фIII) босатылады. Сыртқы механизмнің протромбин активтенуіне дейінгі барлық реакциялары жасуша мембранасының липопротеиндік сынықтарынан тұратын матрицада өтеді.
Ұлпалық тромбопластин – бұл мембраналық фосфогликопротеид. Ол қанның стимулденбеген формалық элементтерінен басқа барлық мембранада болады. Ол өте гидрофобты, сондықтан қалыпты жағдайда қанға түспейді.
Ұлпалық тромбопластиннің теріс зарядталған фосфолипидтеріне уникальді ди-γ-карбоксилглютамин қышқылы қалдығын ұстайтын кальций-бекітуші аймақ есебінен VII фактор байланысады.
Активті фVIIa ұлпалық липидтермен және кальций ионымен комплекс түзеді. Бұл комплекс протеолиттік активтілікке қабілетті және Х фактордың активтенуін жүргізеді.
Активті Xа фактор - протеолиттік активті, сондықтан протромбинді ыдыратып активті қалыпқа (тромбин) әкеледі. (сур.1).
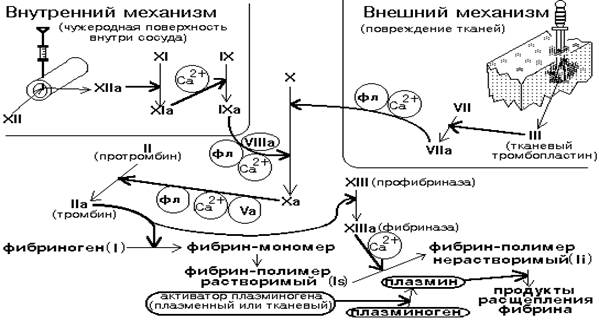 |
Сурет 1. Қан ұюының сыртқы және ішкі механизмдері схемасы
ҰЮДЫҢ ІШКІ МЕХАНИЗМДЕРІНІҢ БАСТАПҚЫ СТАДИЯЛАРЫ
Ұюдың ішкі механизмінің бастапқы стадиялары "жанастырушы фаза" немесе “жанастырушы стадия” деп аталады. Коллагенмен инициацияланады, қалыпты жағдайда ол қан тамырларының ішкі беттерінде болмайды.
Зақымдалған тамырдың беткейінде, құрамында коллаген болатын, қан плазмасының сақшы полижүйесінің үш протеинінен тұратын: - жоғары молекулалы кининогеннен, прекалликреиннен және XII фактордан тұратын комплекс өздігінен жиналады.
Олардың өзара әсерлесуі нәтижесінде XII фактор фXIIа-ға айналады. Ол прекалликреин мен XI факторға фермент ретінде әсер етіп, оларды калликреин және XІа-ға айналдырады.
Калликреин қосымша түрде XII фактор порциясын протеолиттік активтенуге түсіріп үдерісті күшейтеді.
ХІа протеолиттік жолмен IX факторды активтендіреді. Соңғысы антигемофильдік глобулин (парафермент VIIIa және Cа2+) қатысуымен Х факторды Ха факторға айналдырады.
Фосфолипидтік арқалықты ақ тромбаның қалыптасушы жасушалары жасай алады.
Активті Ха тормбоциттердің бетінде протромбинді (плазманың фІІІ және Va-факторы қатысуымен) тромбинге дейін ыдыратады.
Тромбин тромбоциттердің активтелуіне, V, VIII и Х факторлардың агрегациясына, шектеулі протеолизіне мықты кері күшейткіш ықпал береді, бірақ оның негізгі қызметі ф І-ді (фибриногенді) фибринге айналдыру болып табылады.
Ол фибриндік жіптерді тігетін XIII факторды да активтендіреді және ретракцияның орындалуын босатады.
Фибриннің мономерлері спонтанды түрде агрегацияланып ерігіш фибрин-полимердің «жұмсақ» реттелген полимерлік құрылымын түзеді. Нәтижесінде фибрин-полимердің қан компоненттерін тұтуы (ұстап алуы) өтеді – тромб (ұйыма) қалыптасады немесе түзіледі.
Бастапқыда ұйыма борпылдақ және жұмсақ болады, фибрин-полимер молекулалары арасындағы байланыстар әлсіз (ковалентсіз). Бірақ, сонан кейін активті фXIIIa (фибриназа) әсерінен фибрин-полимердің мықты ковалентті байланыстары түзіледі. Глутаминнің карбоксилдік топтары мен лизиннің аминотоптары арасында молекуларалық байланыстар түзіледі: - ерігіш фибрин-полимер ерімейтін фибрин-полимерге айналады.
Фибриннің жіптері түзілген соң олардың жиырылуы (қан ұйымасының ретракциясы) өтеді, оған АТФ жұмсалады.
Тромб түзілуі үдерісі тұрақты түрде сериндік протеиназалардың ингибиторы антитромбин ІІІ арқылы бақыланады. Мұнан басқа, матрицадағы қан ұюының көптеген реакцияларының жүруі қамтамассыз етеді: 1) үдерістің жоғары тиімділігін; 2) үдерістің шоғырлы өтуін – ұю үдерісі тек зақымдалу орнында ғана жүреді (бұл дессиминирленген тамырішілік ұю үдерісін (ДТҰ-синдром) болдырмайды.
Қанның ұю жылдамдығы тек қанның ұю жүйесінің қызметіне ғана тәуелді емес, сонымен бірге қанның ұюын болдырмайтын табиғи антикоагулянттардың болуына да тәуелді.
Фибринолиз – бұл фибрин-полимердің жеке пептидтерге ыдырауы, ол плазмин арқылы катализденеді.
Плазмин - сериндік протеиназа, фибринді, фибриногенді және т.б. гидролиздеуге қабілетті. Плазминнің өзі плазминогеннен плазминоген активаторы әсері арқылы түзіледі. Плазминогеннің ұлпалық активаторы фибринмен жанасқанға дейін активсіз болады. Фибринмен жанасқанда, ол, плазминогенді активтендіруге қабілеттенеді. Фибрин плазминмен гидролизденгенде плазминогеннің активаторы өзінің активтілігін жоғалтады.
Қаннның ұю жүйесі каскадтық реакциялар болып табылады, ал фибринолиз ферменттері бұл каскадтан тыс болады. Мағынасы: - фибринолиз жүйесі және қанның ұю жүйесі біздің ағзамызда тұрақты жұмыс істейді, бірақ өте баяу жылдамдықта. Қалыпты жағдайда, адамда, ұю және фибринолиз үдерістері тепе теңдікте болады. Бұл ағзаның әртүрлі зақымдаушы факторлардың әсеріне қарсы жауап болуына тұрақты дайындығын қамтамассыз етеді. Ағзаның жарақаттануында қанның ұю жүйесінің қызметін өте жылдам күшейтіле алады. Бұл кезде фибринолиз жүйесі плазмин активтілігін тез жоғарлата алмайды, сондықтан ол фибринді гидролиздеуге үлгермейді. Осының нәтижесінде гемостаз атқарылады.
ҰЮҒА ҚАРСЫ ЖҮЙЕ НЕМЕСЕ АНТИКОАГУЛЯНТТАР
Табиғи антикоагулянттар ұлпаларда синтезделеді және қанға түседі де қанның ұю жүйесінің активтенуін тежейді. Оларға гепарин, антитромбин-III және a-2-макроглобулин жатқызылады.
Гепарин кейбір факторлардың активтенуін болдырмайды, бірақ оларға тікелей әсер етпейді. Гепарин антитромбин-III-ті активтендіруге қабілетті. Жоғары теріс заряды болғандықтан ол антитромбин-III-тің катиондық бөлігімен байланысады. Нәтижесінде антитромбин-III-тің конформациясы өзгереді. Ол сериндік протеиназаны инактивтендіруге қабілетті болады.
a-2-Макроглобулин – протеазалардың эндогендік ингибиторы, сонымен бірге қанның ұю жүйесінің қызметіне және фибринолизге (тромбин, плазмин) қатысатын көптеген ферменттердің де.
Протеин С жүйесі п араферменттердің жұмысын бақылайды. Протеин С – бұл гликопротеин, құрамында карбоксиглутамин қышқылы болады, оның синтезі К витаминіне тәуелді. Қанда профермент түрінде болады, тромбинмен активтеледі. Активті протеин С фV және фVIII-ді шектеулі протеолиз жолымен фVa және фVIIIa –ға айналдырады. Қан плазмасында протеин С –ның эндогендік ингибиторы бар.
Қанның ұю жүйесі барлық кезде жұмыс істейді деп есептеледі: бір мезгілде фибриндік ұйыманың түзілуі және ерітілуі өтеді, ол қанның ұю жүйесі мен фибринолиз жүйесінің қызметтері теңесетіндігіне байланысты болады.
Сурет 3-те протеиназалардың активсіз алғызаттары шеңбермен, активті ферменттері боялған қоршаулармен, қосымша факторлары тікбұрыштармен көрсетілген.
ҚАНННЫҢ ФОРМАЛЫҚ ЭЛЕМЕНТТЕРІНІҢ БИОХИМИЯСЫ

