




Под комплементарностью понимают такой тип взаимодействия генов, при котором два гена вместе обусловливают развитие нового признака, отличного от родительских вариантов. Существует не менее трех типов комплементарности:
• доминантные гены различаются по фенотипическому проявлению;
• доминантные гены имеют сходное фенотипическое проявление;
• и доминантные, и рецессивные гены имеют самостоятельное фенотипическое проявление.
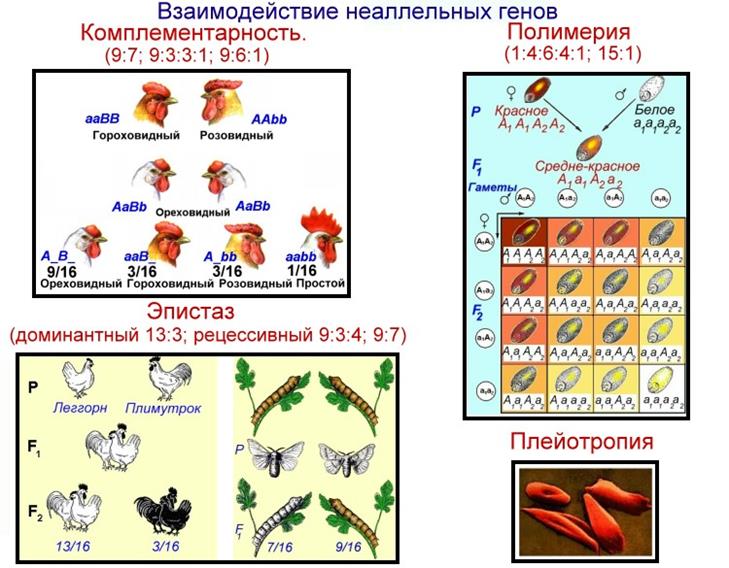 Если доминантные аллели двух генов обусловливают разный фенотип, то в F2, наблюдается расщепление 9:3:3:1. В качестве примера данного типа взаимодействия генов можно привести наследование формы гребня у кур. У гибридов первого поколения доминантные гены А и В дополняют друг друга и вместе обусловливают ореховидную форму гребня, которой не было у родительских форм. При скрещивании гибридов F1: AaBb x AaBb во втором поколении, наряду с ореховидной, розовидной и гороховидной появляется простая форма гребня в соотношении: 9 А_ B_: 3 А_ bb: 3 аа В_: 1 аа bb («_» означает, что аллель в гомологичной хромосоме может быть как доминантным, так и рецессивным).
Если доминантные аллели двух генов обусловливают разный фенотип, то в F2, наблюдается расщепление 9:3:3:1. В качестве примера данного типа взаимодействия генов можно привести наследование формы гребня у кур. У гибридов первого поколения доминантные гены А и В дополняют друг друга и вместе обусловливают ореховидную форму гребня, которой не было у родительских форм. При скрещивании гибридов F1: AaBb x AaBb во втором поколении, наряду с ореховидной, розовидной и гороховидной появляется простая форма гребня в соотношении: 9 А_ B_: 3 А_ bb: 3 аа В_: 1 аа bb («_» означает, что аллель в гомологичной хромосоме может быть как доминантным, так и рецессивным).
При другом варианте комплементарности доминантные аллели двух взаимодействующих генов не имеют собственного фенотипического проявления: новый фенотип у гибридов определяется одновременным присутствием в генотипе двух неаллельных доминантных генов.
Так, у тутового шелкопряда Bombyx mori желтая окраска кокона определяется наличием в генотипе двух доминантных генов-A и В, при наличии в генотипе только одного из этих генов, а также у двойных гомозигот аа bb - окраска кокона белая. Поэтому в F2 у 7/16 коконов окраска белая, а у 9/16 А В_ - желтая.
Расщепление по фенотипу при комплементарном взаимодействии генов:
· 9:7 окраска венчика у гороха (белый, красный)
· 9:6:1 форма плода у тыквы
· 9:3:3:1 форма гребня у кур, окраска плодов перца
При эпистазе происходит подавление действия одного гена другим, не аллельным геном: А > В или А > bb. Гены, подавляющие действие других генов, называют ингибиторами или супрессорами. Они могут быть как доминантными, так и рецессивными. При доминантном эпистазе один доминантный ген подавляет проявление другого доминантного гена.
Возможны два варианта доминантного эпистаза
1.Гомозиготы по рецессивным аллелям фенотипически отличаются от генотипов с доминантными аллелями.
У тыквы Cucurbita pepo окраска плода может быть желтой (А) и зеленой (а). Эта окраска может подавляться доминантным ингибитором I, в результате чего плоды получаются белыми.
Р: IIAA х iiaa
белая зеленая
F1: IiАа
белая
F2: 9/16 I_A_: 3/16 I_ аа: 3/l6 iiA_: 1/16 ii аа
белые(12) желтые(3) зеленые(1)
В описанном и аналогичных случаях в F2 имеет место расщепление по фенотипу 12:3:1.
2. Гомозиготы по рецессивным аллелям iiaa не отличаются по фенотипу от I_А_ и от I_ аа.
Например, у кукурузы Zea mays окраска зерна может быть пурпурной (А) и белой (а).
У 9/16 растений I_А_ при наличии доминантного ингибитора I пигмент не синтезируется. У 3/16 растений с генотипом I_аа и у 1/16 iiаа отсутствует ген пурпурной окраски, поэтому зерна в початках кукурузы также белые. И только у 3/16 iiА_ окраска зерен пурпурная, поскольку в генотипе есть ген окраски, а ингибитор представлен рецессивным аллелем.
Таким образом, в F2 соотношение растений с окрашенными и неокрашенными зернами 13:3.
Р: IIАА х iiaa
белая белая
F1: IiАа
белая
F2: 9/16 I_A_: 3/16 I_аа: 1/16 iiaa: 3/16 iiА_
белые (13) пурпурные (3)
При рецессивном эпистазе рецессивный аллель одного гена подавляет действие неаллельного доминантного гена aa > В_, а между доминантными генами наблюдается комплементарность. Например, у льна Linum usitatissimum аллель А определяет окрашенный венчик, аа — неокрашенный (белый), В - голубой, bb - розовый. По-видимому, ген А необходим для синтеза предшественника пигмента, без которого ни голубой, ни розовый пигменты не образуются. Гетерозиготные растения А_В_ имеют голубую окраску венчика (комплементарность доминантных генов), AaBb- розовую, тогда как рецессивные аллели гена а в гомозиготном состоянии подавляют синтез как голубого пигмента в генотипах ааВВ и ааВЬ, так и розового пигмента у ааbb (рецессивный эпистаз).
Вследствие двух типов взаимодействия генов у льна в F2 наблюдается расщепление 9:3:4.
Р: ААbb х ааВВ
розовый белый
F1 AaBb
голубой
F2: 9/I6A_B_: 3/16A_bb: 3/16 аа В_: 1/16 аа bb
голубые (9) розовые (3) белые (4)
Полимерия, полигения, обусловленность одного сложного признака многими неаллельными генами, действие которых суммируется в признаке. Такие гены называются полигенами. В условиях неоднородной внешней среды полимерия приводит к непрерывной, или количественной, изменчивости признака в популяции. Большинство признаков относится к количественным, например размеры и вес особей, их окраска, иногда устойчивость к заболеваниям, многие хозяйственные полезные признаки с.-х. животных (удой и жирномолочность у коров, настриг и окраска шерсти у овец, яйценоскость и размеры яиц у кур и т.д.), с.х. растений (сахаристость у сахарной свеклы, количество белка и клейковины в зерне пшеницы и т.д.). Полимерия была открыта в 1909 шведским учёным Г. Нильсоном-Эле, изучавшим наследование окраски зёрен у пшеницы путём анализа расщеплений этого признака.

