Нуклеиновые кислоты – это линейные неразветвленные гетерополимеры, мономерами которых являются нуклеотиды, связанные фосфодиэфирными связями.
Существует два типа нуклеиновых кислот: ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота) (рис.2).
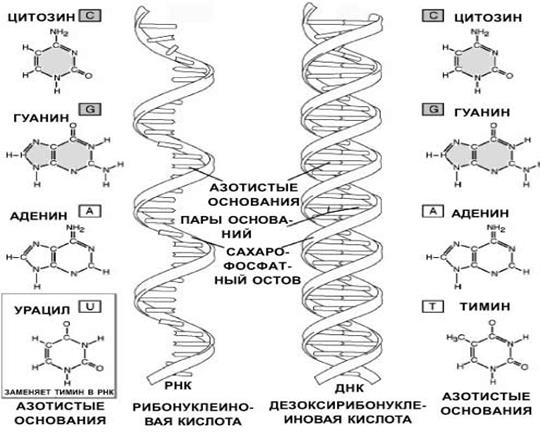 |
Нуклеотиды – это органические вещества, молекулы которых состоят из остатка пентозы (рибозы или дезоксирибозы), к которому ковалентно присоединены остаток фосфорной кислоты и азотистое основание.
Рис.2. Строение нуклеиновых кислот
Азотистые основания в составе нуклеотидов делятся на две группы: пуриновые (аденин и гуанин) и пиримидиновые (цитозин, тимин и урацил).
Дезоксирибонуклеотиды включают в свой состав дезоксирибозу и одно из азотистых оснований: аденин (А), гуанин (Г), тимин (Т), цитозин (Ц).
Рибонуклеотиды включают в свой состав рибозу и одно из азотистых оснований: аденин (А), гуанин (Г), урацил (У), цитозин (Ц).
ДНК – самая большая молекула в клетке. Она намного больше белков и РНК.
Каждая хромосома - это одна молекула ДНК.
ДНК – это биополимер с молекулярной массой 10 -100 млн., мономерами которого являются 4 типа нуклеотидов. В последовательности ее нуклеотидов записана вся наследственная программа организма
Функция ДНК — хранение и передача наследственной информации.
В основе построения молекулы ДНК лежит правила Чаргаффа:
1. Сумма пуриновых нуклеотидов (Пур) равна сумме пиримидиновых нуклеотидов (Пир) (Пур = Пир, или Пур/Пир=1).
2. Молярное содержание аденина равно молярному содержанию тимина (А = Т, или А/Т= 1).
3. Молярное содержание гуанина равно молярному содержанию цитозина (Г = Ц, или Г/Ц=1).
4. Количество аденина и цитозина равно количеству гуанина и тимина (А + Ц=Г + Т, или А + Ц/Г + Т=1).
5. В ДНК из различных источников неодинаково соотношение нуклеотидов: у одних преобладает содержание аденина над гуанином, тимина над цитозином (А + Т>Г + Ц) — это так называемый АТ-тип ДНК, у других преобладают гуанин и цитозин над аденином и тимином (Г + Ц>А + Т), это — ГЦ-тип ДНК. Таким образом, ДНК из различных источников отличается по нуклеотидному составу, что выражается различной величиной отношения Г + Ц/А + Т. В связи с этим Э. Чаргафф выдвинул положение о видовой специфичности ДНК по нуклеотидному составу.
Объяснение правилам Чаргаффа дали Уотсон и Крик.
ДНК – это 2 цепочки, соединенные по принципу комплементарности.
Существует несколько типов ДНК: А, В, С, Z – формы.
Таблица 2. Формы ДНК
| Параметры | B-форма | А-форма | С-форма | Z-форма |
| спираль | правозакручена | правозакручена | правозакручена | левозакручена |
| ед. повтора | 1 пн | 1пн | 1пн | 2пн |
| пн в обороте | 10,4 | 10,7 | 9.3 | |
| диаметр | 23,7А | 25,5А | 18,4А | |
| вращение/пн | 35,9 | 33,6 | 38,7 | 60/2 |
| наклон пн к оси | -1,2 | +19 | -9 | |
| раст. м-у пн вдоль оси | 0.332 nm | 0.23 nm | 0.38 nm | |
| длина оборота | 34А | 28А | 31А | 34,4А |
Репликация ДНК — процесс самоудвоения, главное свойство молекулы ДНК.
Ø Репликация относится к категории реакций матричного синтеза, идет с участием ферментов.
Ø Репликация протекает сходно у прокариот и эукариот.
Ø Скорость синтеза ДНК у прокариот на порядок выше (1000 нуклеотидов в секунду), чем у эукариот (100 нуклеотидов в секунду).
Ø Репликация начинается одновременно в нескольких участках молекулы ДНК.
Ø Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
Ø Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Принципы репликации
1. Комплементарность – в соответствии с правилами Чаргаффа
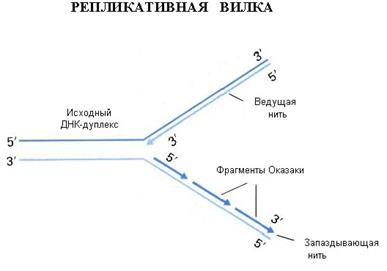
2. Антипараллельность цепей ДНК – противоположная направленность двух нитей двойной спирали ДНК; одна нить имеет направление от 5' к 3', другая - от 3' к 5'.
3. Полуконсервативность - в результате репликации образуются две двойные дочерние спирали, каждая из которых сохраняет (консервирует) в неизменном виде одну из половин материнской ДНК. Вторые цепи дочерних молекул синтезируются из нуклеотидов заново по принципу комплементарности к нитям материнской ДНК.
4. Униполярность – ферменты, синтезирующие новые нити ДНК, называемые ДНК-полимеразами, могут передвигаться вдоль матричных цепей лишь в одном направлении - от их 3'-концов к 5'-концам. При этом синтез комплементарных нитей всегда ведется в 5'→ 3' направлении, то есть униполярно. Поэтому в процессе репликации одновременный синтез новых цепей идет антипараллельно.
5. Прерывистость – для того, чтобы новые нити ДНК были построены по принципу комплементарности, двойная спираль должна быть раскручена и родительские цепи вытянуты. Но раскручивание спиралей, состоящих из многих миллионов пар нуклеотидов, сопряжено со столь значительным количеством вращений и такими энергетическими затратами, которые невозможны в условиях клетки. Поэтому репликация начинается одновременно в нескольких местах молекулы ДНК. Участок между двумя точками, в которых начинается синтез дочерних цепей, называется репликоном. Он является единицей репликации. В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку - ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке несколько ферментов одновременно ведут синтез ДНК в виде фрагментов по 1000 нуклеотидов. Эти фрагменты имеют противоположную направленность. Когда вилка перемещается, синтезированные фрагменты, комплементарные одной цепи, сшиваются друг с другом, образуя растущие дочерние цепи. Такой механизм синтеза новых цепей ДНК фрагментами называется прерывистым.
6. Потребность в затравке – ДНК-полимераза не способна начать синтез. Она может лишь наращивать уже имеющуюся полинуклеотидную нить. Откуда же берётся начальный 5'-концевой участок растущей цепи? Его синтезирует особая РНК-полимераза, называемая праймазой (от англ. primer - затравка). Размер рибонуклеотидной затравки невелик (< 20 нуклеотидов). Выполнившая свою функцию РНК-затравка удаляется специальным ферментом.