




В. М. Смирнов, В. И. Дубровский
ФИЗИОЛОГИЯ
ФИЗИЧЕСКОГО ВОСПИТАНИЯ И СПОРТА
Рекомендовано Государственным комитетом
Российской Федерации по физической культуре,
спорту и туризму в качестве учебника
для средних и высших учебных заведений
по физической культуре

ББК 75.0я73 С50
Рецензенты:
доктор медицинских наук, профессор В. Н. Яковлев;
доктор медицинских наук, профессор В. А. Правдивцев;
доктор медицинских наук, профессор В. И. Торшин
Художественное оформление Н. М. Замешаева
Смирнов В. М., Дубровский В. И.
С50 \ Физиология физического воспитания и спорта: Учеб. для студ. сред, и высш. учебных заведений. — М.: Изд-во ВЛАДОС-ПРЕСС, 2002. — 608 с: ил.
15Г^ 5-305-00034-3.
Учебник написан в соответствии с новой программой изучения «Физиологии физического воспитания и спорта» в различных учебных заведениях. Большое внимание уделено физиологии ЦНС и нервно-мышечной системы, терморегуляции и акклиматизации, биоритмам, гормональной регуляции при физической деятельности, другим вопросам, касающимся физической работы, физиологическим изменениям в функциональных системах и органах под влиянием физических нагрузок; дана характеристика состояния организма при занятиях различными видами спорта и оздоровительной физкультурой.
Учебник рассчитан на студентов колледжей, факультетов физической культуры университетов и институтов физической культуры, медицинских вузов, а также тренеров, спортивных врачей и других специалистов.
ББК 75.0я73
© Смирнов В. М., Дубровский В. И., 2002
© «Издательство ВЛАДОС-ПРЕСС», 2002
© Серийное оформление обложки.
15ВЫ 5-305-00034-3 «Издательство ВЛАДОС-ПРЕСС», 2002
ПРЕДИСЛОВИЕ
Учебник «Физиология физического воспитания и спорта» профессора В. М. Смирнова и профессора В. И. Дубровского написан согласно новой программе и состоит из трех частей: физиология зрелого организма, особенности физиологии детей и подростков и собственно спортивная физиология.
Часть I учебника оформлена в традиционном для учебников по физиологии стиле - по отдельным системам - и состоит из трех разделов.
Первый раздел «Физиология возбудимых тканей» содержит материалы по общей физиологии возбудимых тканей, физиологии сенсорных рецепторов, нервных волокон, синапсов, общей и частной физиологии ЦНС, эндокринным железам. Несмотря на краткость изложения этого материала, авторы смогли четко и понятно представить классические и современные данные по физиологии и биофизике возбудимых клеток. Особое внимание уделено вопросам физиологии движений, что вполне обосновано целевым назначением учебника. Следует также отметить, что это ключевой раздел для глубокого понимания механизмов деятельности всех органов и систем организма.
Второй раздел посвящен физиологии внутренних органов и систем организма - это система крови, дыхания, сердечно-сосудистая система, пищеварительная, обмен веществ и энергии, терморегуляция, выделительная система. Большое внимание в этом, как и в других разделах учебника, уделено терминам, по которым еще не сложилось единого мнения. Авторы в ряде случаев выступают как
новаторы, причем убедительно обосновывают свою оригинальную точку зрения, не навязывая ее читателю. Напротив, читателю предоставляется возможность творческого решения дискуссионного вопроса.
Третий раздел «Интегративная деятельность организма» посвящен физиологии сенсорных систем и высшей нервной деятельности. Здесь дана оригинальная классификация анализаторов, четко изложены механизмы кодирования информации в различных отделах анализаторов, особое внимание уделено кодированию раздражителей в периферическом - рецепторном и в корковом отделах анализатора (кодирование качественной характеристики действующего на рецепторы раздражителя, количественной - сила раздражителя, кодирование пространства и времени действия раздражителя). Авторы изложили современные представления о механизмах кодирования в корковом конце анализатора - о механизмах узнавания известного предмета, явления и формирования образа, впервые встречаемого предмета и явления. Достаточно глубоко, хотя и компактно, написан раздел частной физиологии анализаторов, оригинально представлен материал по физиологии зрительного анализатора. Дана обоснованная, облегчающая усвоение материала модификация классификации различных вариантов научения, сформулированы оригинальные определения - что такое психическая деятельность, первая и вторая сигнальные системы, четко изложены устоявшиеся представления о механизмах ВНД.
В части II учебника детально изложены материалы, касающиеся особенностей физиологии детей и подростков, начиная с периода новорожденности до юношеского возраста, рассмотрена проблема акселерации и ретардации развития детей. Показано созревание различных функциональных систем организма, в частности, особенности опорно-двигательного аппарата у детей, особенности терморегуляции и обмена веществ, переход от развивающегося организма к зрелому.
В части III учебника отражены современные представления о влиянии физической культуры и спорта на физиологические сис-
темы, органы и ткани организма как во время тренировки, так и в соревновательной деятельности - каким образом органы и физиологические системы обеспечивают физические нагрузки. Впервые отмечены факторы негативного влияния спорта на организм спортсмена. Представлены данные о влиянии гипоксии (тренировки в горах), жаркого и влажного климата на тренирующихся спортсменов.
В учебник введен раздел адаптации (акклиматизации) спортсменов к условиям смены временных поясов и участия спортсменов в международных соревнованиях в различных климатических условиях. В частности, представлены данные о влиянии климата и биологических ритмов на спортивные результаты во время тренировок и участия в международных соревнованиях российских спортсменов в США, Канаде, Перу, Камбодже и других странах. Описаны особенности реакций кардиореспираторной системы, метаболизма, гомеостазиса на физические нагрузки. Отмечена динамика гомео-стазиса при продолжительной акклиматизации спортсменов и при физических нагрузках в период подготовки к ответственным соревнованиям.
Сделан акцент на особенности метаболизма тканей, органов и функциональных систем в условиях адаптации спортсменов к чрезмерным физическим нагрузкам. Представлены данные о физиологических реакциях у юных спортсменов, женщин и лиц пожилого возраста на влияние физических тренировок.
Особое внимание уделено описанию физиологической характеристики таких качеств,,как выносливость, скорость, сила; физиологической характеристике различных видов спорта (циклические, ациклические, единоборства и др.); аэробным и анаэробным фазам при различных физических нагрузках и внешних факторах.
Книга прекрасно иллюстрирована схемами, таблицами, рисунками. Иллюстрации, несомненно, будут способствовать повышению эффективности преподавания и усвоения материала. В иллюстрациях отражены представления о некоторых процессах, протекающих на тканевом и клеточном уровнях.

|
Поскольку физиология является одной из основополагающих наук, ее изучение необходимо будущему тренеру, учителю физкультуры, спортивному врачу и другим специалистам, которые занимаются физической культурой и спортом. Изучение физиологии физического воспитания необходимо в связи с массовым развитием физической культуры и спорта высших достижений; знание физиологии необходимо каждому человеку для успешных занятий физической культурой, и особенно лицам, имеющим отклонения в состоянии здоровья.
Академик РАМН, директор НИИ
физиологии им. П. К. Анохина,
заведующий кафедрой физиологии
Академии им. И. М. Сеченова,
профессор К. В. СУДАКОВ
Раздел I
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ. РЕГУЛЯЦИЯ ФУНКЦИЙ ОРГАНИЗМА
Глава 1
ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Возбудимыми являются ткани, способные генерировать потенциал действия (возбуждаться), - это нервная и мышечная ткани.
Сущность процесса возбуждения заключается в следующем. Все клетки организма имеют электрический заряд, обеспечиваемый неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различная концентрация анионов и катионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для разных ионов и работы ионных насосов. При действии раздражителя на клетку возбудимой ткани изменяется проницаемость ее мембраны, вследствие чего ионы быстро перемещаются согласно электрохимическому градиенту (совокупность концентрационного и электрического градиентов), - это и есть процесс возбуждения. Его основой является потенциал покоя.
ПОТЕНЦИАЛ ПОКОЯ
А. Общая характеристика и непосредственная причина формирования. Потенциал покоя (ПП) - относительно стабильная разность между величинами электрических потенциалов внутри и вне клетки в покое. Его величина обычно варьирует в пределах 30-90 мВ (в волокнах скелетной мышцы - 60-90 мВ, в нервных клетках -50-80 мВ, в гладких мышцах -30-70 мВ, в сердечной мышце -80-90 мВ). При регистрации ПП луч осциллографа во время прокола мембраны клетки микроэлектродом скачком отклоняется и показывает отрицательный заряд внутри клетки (рис. 1.1).

ПП играет исключительно важную роль в жизнедеятельности самой клетки и организма в целом. В частности, он составляет основу для переработки информации нервной клеткой, обеспечи-* вает регуляцию деятельности внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения в мышце. Нарушение процессов возбуждения в кардио-миоцитах ведет к остановке сердца. Согласно мембранно-ионной теории (Бернштейн, Ходжкин, Хаксли, Катц, 1902-1952), непосредственной причиной формирования ПП является неодинаковая концентрация анионов и катионов внутри и вне клетки.
В нервных и мышечных клетках концентрация К+ внутри клетки в 30-40 раз больше, чем вне клетки; концентрация №+ вне клетки в 10-12 раз больше, нежели внутри клетки; ионов С1~ вне клетки в 15-20 раз больше, чем внутри клетки. В клетке имеется небольшое количество ионов М§2+. Кальций в свободном состоянии находится в основном вне клетки. Он содержится также в эн-доплазматическом ретикулуме; в гиалоплазме его очень мало. Это обусловливается отчасти активным транспортом Са2+ наружу через клеточную мембрану, отчасти поглощением его эндоплазмати-ческим ретикулумом - это резервуар для Са2+- и другими органел-лами, например, митохондриями, связыванием Са2+ цитратом, глютаматом.
В клетке находятся также крупномолекулярные анионы - главным образом это отрицательно заряженные белковые молекулы,
например, глютамат, аспартат, а также органические фосфаты. Различные ионы распределены неравномерно по обе стороны клеточной мембраны, во-первых, вследствие неодинаковой проницаемости клеточной мембраны для различных ионов, во-вторых, в результате работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки концентрационному и электрическому градиентам.
Б. Терминология и характеристика клеточной мембраны.
Проницаемость клеточной мембраны - это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам диффузии и фильтрации. Проницаемость клеточной мембраны (она представляет собой липопротеиновую оболочку) определяется следующими факторами: 1) наличием в составе мембраны различных ионных каналов - управляемых (с воротным механизмом) и неуправляемых (каналов утечки); 2) размерами каналов и размерами частиц; 3) растворимостью частиц в мембране (клеточная мембрана проницаема для растворимых в ней липи-дов и непроницаема для пептидов).
Проводимость - это способность заряженных частиц (ионов) проходить через клеточную мембрану согласно электрохимическому градиенту. Проводимость иона зависит от его электрохимического градиента и от проницаемости мембраны - чем они больше, тем лучше проводимость иона через мембрану. Перемещения ионов в клетку и из клетки, согласно концентрационному и электрическому градиентам в состоянии покоя клетки, осуществляются преимущественно через неуправляемые (без воротного механизма) каналы, их называют также каналами утечки. Неуправляемые каналы всегда открыты, они практически не меняют своей пропускной способности при электрическом воздействии на клеточную мембрану и ее возбуждении. Неуправляемые каналы подразделяются на ионоселективные каналы (например, калиевые медленные неуправляемые каналы) и иононеселективные каналы. Последние пропускают различные ионы - К+, Ыа+, СГ.
В. Роль проницаемости клеточной мембраны и различных ионов в формировании ПП. №+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам диффузии, при этом К+ из клетки выходит в значительно большем количестве, чем входит Ыа+ в клетку, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости для Ыа+. Органические анионы из-за своих больших размеров не могут выходить из клетки, поэтому внутри клетки в состоянии покоя отрицательных ионов оказывается больше, чем положительных. Однако превышение абсолютного числа анионов над числом катио-
нов в клетке чрезвычайно мало. Но этого различия достаточно для создания разности электрических потенциалов внутри и вне клетки. Главным ионом, обеспечивающим формирование ПП, является ион К+ (рис. 1.2). Об этом свидетельствуют результаты опыта с перфузией внутреннего содержимого гигантского аксона кальмара солевыми растворами. При уменьшении концентрации К+ в перфузате ПП уменьшается, при увеличении концентрации К+ ПП увеличивается. В покоящейся клетке устанавливается динамическое равновесие между числом выходящих из клетки и входящих в клетку ионов К+. Электрический и концентрационный градиенты противодействуют друг другу - согласно концентрационному градиенту К+ стремится выйти из клетки, отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточ-
ной мембраны препятствуют этому. Когда концентрационный и электрический градиенты уравновесятся, число выходящих из клетки ионов К+ сравняется с числом входящих в клетку ионов К+. В этом случае на клеточной мембране устанавливается так называемый равновесный калиевый потенциал.
Равновесный потенциал для любого иона можно рассчитать по формуле Нернста. Концентрация положительно заряженного иона, находящегося снаружи, в формуле Нернста располагается в числителе, иона, находящегося внутри клетки, - в знаменателе. Для отрицательно заряженных ионов расположение противоположное:


|
где Е(оп - потенциал, создаваемый данным ионом; К - газовая постоянная (8,31 Дм); Т - абсолютная температура (273+37°С); 2 -валентность иона; Р - постоянная Фарадея (9,65 104); [юп]\ - концентрация иона внутри клетки ОпзШе); [юп]о - концентрация иона во внешней среде клетки (оиЫае).
Вклад Л^а+ и СГ в создание ПП. Проницаемость клеточной мембраны в покое для Ыа+ очень низкая - намного ниже, чем для К+, тем не менее она имеет место, поэтому ионы №+, согласно концентрационному и электрическому градиентам, стремятся и в небольшом количестве проходят внутрь клетки. Это ведет к уменьшению ПП, так как на внешней поверхности клеточной мембраны суммарное число положительно заряженных ионов уменьшается, хотя и незначительно, а часть отрицательных ионов внутри клетки нейтрализуется входящими в клетку положительно заряженными ионами №+. Что касается СГ, его влияние на величину ПП противоположно влиянию №+ и зависит от проницаемости клеточной мембраны для СГ (она в 2 раза ниже, чем для К+). Дело в том, что СГ, согласно концентрационному градиенту, стремится и проходит в клетку, что ведет к увеличению ПП. Концентрации ионов К+ и СГ близки между собой. Но СГ находится в основном вне клетки, а К+ - внутри клетки. Препятствует входу СГ в клетку электрический градиент, поскольку заряд внутри клетки отрицательный, как и заряд СГ. Наступает равновесие сил концентрационного градиента, способствующего входу СГ в клетку, и электрического градиента, препятствующего входу СГ в клетку.
Г. Определенную роль в формировании ПП играют поверхностные заряды самой клеточной мембраны и ионы Са2+. Наружная и внутренняя поверхности клеточной мембраны несут собственные электрические заряды, преимущественно с от-
рицательным знаком. Это полярные молекулы клеточной мембраны: гликолипиды, фосфолипиды, гликопротеиды. Фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, уменьшают ПП. Фиксированные внутренние отрицательные заряды клеточной мембраны, напротив, суммируясь с анионами внутри клетки, увеличивают ПП. Роль ионов Са2+ в формировании ПП заключается в том, что они взаимодействуют с наружными отрицательными фиксированными зарядами мембраны клетки и отрицательными карбоксильными группами интерстиция и нейтрализуют их, что ведет к увеличению и стабилизации ПП.
Таким образом, ПП - это алгебраическая сумма не только всех зарядов ионов вне и внутри клетки, но также алгебраическая сумма отрицательных внешних и внутренних поверхностных зарядов самой мембраны.
При проведении измерений потенциал окружающей клетку среды принимают за величину, равную нулю. Относительно нулевого потенциала внешней среды потенциал внутренней среды клетки, как отмечалось выше, составляет величину порядка 60-90 мВ. Повреждение клетки приводит к повышению проницаемости клеточных мембран, в результате чего различие проницаемости для,, К+ и Ка+ уменьшается, ПП при этом снижается.
Д. Роль ионных насосов в формировании ПП. В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки постепенно должна выравниваться. Однако, несмотря на постоянную диффузию ионов (утечку ионов), ПП клеток остается на одном уровне. Следовательно, кроме собственно ионных механизмов формирования ПП, связанных с различной проницаемостью клеточной мембраны, имеется активный механизм поддержания градиентов концентрации различных ионов внутри и вне клетки. Им являются ионные насосы, в частности Ыа/К-насос (помпа).
Ионный насос - это транспортная система, обеспечивающая перенос иона с непосредственной затратой энергии вопреки концентрационному и электрическому градиентам. Если заблокировать освобождение энергии, например динитрофенолом, в течение 1 ч выведение Ыа+ из клетки сократится примерно в 100 раз. Как выяснилось, выведение №+ сопряжено с транспортом К+, что можно продемонстрировать при удалении К+ из наружного раствора. Если К+ на наружной стороне мембраны нет, работа насоса блокируется, перенос Ыа+ из клетки в этом случае падает, составляя примерно 30% от нормального уровня. Сопряженность транспорта Ыа+ и К+ уменьшает расход энергии примерно в 2 раза по сравне-
нию с той, которая потребовалась бы при несопряженном транспорте. В целом, траты энергии на активный транспорт веществ огромны - лишь Ыа /К- насос потребляет 1 / 3 всей энергии, расходуемой организмом в покое. За 1 с один Ыа/К-насос (одна молекула белка) переносит 150-600 ионов №+. Накопление №+ в клетке стимулирует работу №/К-насоса, уменьшение Ыа+ в клетке снижает его активность, поскольку снижается вероятность контакта ионов с соответствующим переносчиком. В результате сопряженного транспорта Ыа+ и К+ поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ обеспечивает один цикл работы №/К-насо-са - перенос трех ионов Ыа+ за пределы клетки и двух ионов К+ внутрь клетки. Асимметричный перенос ионов №/К- насосом поддерживает избыток положительно заряженных частиц на наружной поверхности клеточной мембраны и отрицательных зарядов внутри клетки, что позволяет считать № / К-насос структурой электрогенной, дополнительно увеличивающей ПП примерно на 5-10 мВ (в среднем около 10% у разных возбудимых клеток -у одних больше, у других меньше). Данный факт свидетельствует о том, что решающим фактором в формировании ПП является селективная проницаемость клеточной мембраны для разных ионов. Если уравнять проницаемость клеточной мембраны для всех ионов, то ПП будет составлять только 5-10 мВ - за счет работы Ыа/К-помпы.
Нормальная величина ПП является необходимым условием возникновения процесса возбуждения клетки, т. е. возникновения и
аспространения потенциала действия, инициирующего специфи-
ескую деятельность клетки.
ПОТЕНЦИАЛ ДЕЙСТВИЯ
Потенциал действия (ПД) - это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала вследствие перемещения ионов в клетку и из клетки и способный распространяться без декремента (без затухания). ПД обеспечивает передачу сигналов между нервными клетками, нервными центрами и рабочими органами; в мышцах ПД обеспечивает процесс электромеханического сопряжения.
А. Характеристика потенциала действия (ПД). Схематично ПД представлен на рис. 1.3. Величина ПД колеблется в пределах 80-130 мВ, длительность пика ПД нервного волокна 0,5-1 мс, волокна скелетной мышцы - до 10 мс с учетом замедле-
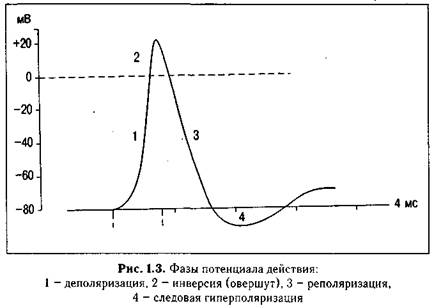
ния деполяризации в конце ее. Длительность ПД сердечной мышцы,, 300-400 мс. Амплитуда ПД не зависит от силы раздражения - она всегда максимальна для данной клетки в конкретных условиях: ПД подчиняется закону «все или ничего», но не подчиняется закону силовых отношений - закону силы. ПД либо совсем не возникает при раздражении клетки, если оно мало, либо возникает и достигает максимальной величины, если раздражение является пороговым или сверхпороговым. Следует отметить, что слабое (подпороговое) раздражение может вызвать локальный потенциал. Он подчиняется закону силы - с увеличением силы стимула величина его возрастает. В составе ПД различают четыре фазы: 1 — деполяризация, т. е. исчезновение заряда клетки - уменьшение мембранного потенциала до нуля; 2 — инверсия, т. е. изменение заряда клетки на противоположный, когда внутренняя сторона мембраны клетки заряжается положительно, а внешняя - отрицательно (лат. шуегзю - переворачивание); 3 — реполяризация, т. е. восстановление исходного заряда клетки, когда внутренняя поверхность клеточной мембраны снова заряжается отрицательно, а наружная -положительно; 4 - следовая гиперполяризация.
Б. Механизм возникновения ПД. Если действие раздражителя на клеточную мембрану приводит к началу развития ПД, далее сам процесс развития ПД вызывает фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое
движение №+ в клетку, а К+ - из клетки. Это наиболее часто встречаемый вариант возникновения ПД. Величина мембранного потенциала при этом сначала уменьшается, а затем снова восстанавливается до исходного уровня. На экране осциллографа отмеченные изменения мембранного потенциала предстают в виде пикового потенциала - ПД. Он возникает вследствие накопленных и поддерживаемых ионными насосами градиентов концентраций ионов внутри и вне клетки, т.е. за счет потенциальной энергии в виде электрохимических градиентов ионов. Если заблокировать процесс выработки энергии, потенциалы действия некоторый период времени будут возникать. Но после исчезновения градиентов концентраций ионов (устранения потенциальной энергии) клетка генерировать ПД не будет. Рассмотрим фазы ПД.
1. Фаза деполяризации (см. рис. 1.3 - 1). При действии деполяризующего раздражителя на клетку (медиатор, электрический ток) начальная частичная деполяризация клеточной мембраны происходит без изменения ее проницаемости для ионов. Когда деполяризация достигает примерно 50% пороговой величины (50% порогового потенциала), начинает повышаться проницаемость мембраны клетки для Ыа+, причем в первый момент сравнительно медленно. Естественно, что скорость входа Ыа+ в клетку при этом невелика. В этот период, как и во время всей первой фазы (деполяризации), движущей силой, обеспечивающей вход Гч!а+ в клетку, являются концентрационный и электрический градиенты. Напомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются друг к другу), а концентрация №+ вне клетки в 10-12 раз больше, чем внутри клетки. Условием, обеспечивающим вход №+ в клетку, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма Ыа-каналов (в некоторых клетках, например, в кардиомиоцитах, в волокнах гладкой мышцы, важную роль в возникновении ПД играют и управляемые каналы для Са2+).
Когда деполяризация клетки достигает критической величины (Е, критический уровень деполяризации - КУД), которая обычно составляет 50 мВ (возможны и другие величины), проницаемость мембраны для Ыа* резко возрастает - открывается большое число потенциалзависимых ворот Ыа-каналов - и Ыа+ лавиной устремляется в клетку. В результате интенсивного тока Ыа+ внутрь клетки процесс деполяризации проходит очень быстро. Развивающаяся деполяризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, естественно, проводимости Ыа+ -открываются все новые и новые ворота №-каналов, что придает току Ыа+ в клетку характер регенеративного процесса. В итоге ПП
исчезает, становится равным нулю. Фаза деполяризации на этом заканчивается.
2. Фаза инверсии. После исчезновения ПП вход Ыа+ в клетку продолжается, поэтому число положительных ионов в клетке превосходит число отрицательных ионов, заряд внутри клетки становится положительным, снаружи - отрицательным. Процесс перезарядки мембраны представляет собой вторую фазу потенциала действия - фазу инверсии (рис. 1.3 - 2). Теперь электрический градиент препятствует входу Ыа+ внутрь клетки (положительные заряды отталкиваются друг от друга), Ыа-проводимость снижается. Тем не менее, некоторый период времени (доли миллисекунды) №+ продолжает входить в клетку — об этом свидетельствует продолжающееся нарастание ПД. Это означает, что концентрационный градиент, обеспечивающий движение №+ в клетку, сильнее электрического, препятствующего входу Ыа+ в клетку. Во время деполяризации мембраны увеличивается проницаемость ее и для Са2+, он также идет в клетку, но в нервных волокнах, нейронах и в клетках скелетной мускулатуры роль Са2+ в развитии ПД мал.а. В клетках гладкой мышцы и миокарда его роль существенна. Таким образом, вся восходящая часть пика ПД в большинстве случаев обеспечивается в основном входом №+ в клетку.
Примерно через 0,5-1 мс и более после начала деполяризации (это время зависит от вида клетки) рост ПД прекращается вследствие закрытия ворот натриевых каналов и открытия ворот К-кана-лов, т. е. увеличения проницаемости для К+ и резкого возрастания выхода его из клетки (см. рис. 1.3 - 2). Препятствуют также росту пика ПД электрический градиент Ыа+ (клетка внутри в этот момент заряжена положительно), а также выход К+ из клетки по каналам утечки. Поскольку К+ находится преимущественно внутри клетки, он, согласно концентрационному градиенту, быстро выходит из клетки после открытия ворот К+-каналов, вследствие чего уменьшается число положительно заряженных ионов в клетке. Заряд клетки снова начинает уменьшаться. В фазу инверсии выходу К+ из клетки способствует также и электрический градиент. К+ выталкивается положительным зарядом из клетки и притягивается отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки (до конца фазы инверсии - рис. 1.3-2, пунктирная линия), когда начинается следующая фаза ПД - фаза реполяризации. Калий выходит из клетки не только по управляемым каналам, ворота которых открыты, но и по неуправляемым - каналам утечки, что несколько замедляет ход восходящей части ПД и ускоряет ход нисходящей составляющей ПД.
Таким образом, изменение мембранного потенциала покоя ведет к последовательному открытию и закрытию электроуправляе-мых ворот ионных каналов и движению ионов согласно электрохимическому градиенту - возникновению ПД. Все фазы являются регенеративными - необходимо только достичь критического уровня деполяризации, далее ПД развивается за счет потенциальной энергии клетки в виде электрохимических градиентов, т. е. вторично активно.
Амплитуда ПД складывается из величины ПП (мембранного потенциала покоящейся клетки) и величины фазы инверсии, составляющей у разных клеток 10-50 мВ. Если мембранный потенциал покоящейся клетки мал, амплитуда ПД этой клетки небольшая.
3. Фаза реполяризации (рис. 1.3-3) связана с тем, что проницаемость клеточной мембраны для К+ все еще высока (ворота калиевых каналов открыты), К+ продолжает быстро выходить из клетки, согласно концентрационному градиенту. Поскольку клетка теперь уже снова внутри имеет отрицательный заряд, а снаружи - положительный (см. рис. 1.3 - 3), электрический градиент препятствует выходу К+ из клетки, что снижает его проводимость, хотя он продолжает выходить. Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее электрического градиента. Вся нисходящая часть пика ПД обусловлена выходом К+ из клетки. Нередко в конце ПД наблюдается замедление реполяризации, что объясняется уменьшением проницаемости клеточной мембраны для К+ и замедлением выхода его из клетки из-за частичного закрытия ворот К-каналов. Вторая причина замедления тока К+ из клетки связана с возрастанием положительного потенциала наружной поверхности клетки и формированием противоположно направленного электрического градиента.
Таким образом, главную роль в возникновении ПД играет Ыа+, входящий в клетку при повышении проницаемости клеточной мембраны и обеспечивающий всю восходящую часть пика ПД. При замене Ма+ в среде на другой ион, например холин, ПД в нервной и мышечной клетках скелетной мускулатуры не возникает. Однако проницаемость мембраны для К+ тоже играет важную роль. Если повышение проницаемости для К+ предотвратить тетраэтиламмонием, мембрана после ее деполяризации реполя-ризуется гораздо медленнее, только за счет медленных неуправляемых каналов (каналов утечки ионов), через которые К+ будет выходить из клетки.

|
Роль Са2+ в возникновении ПД в нервных и мышечных клетках скелетной мускулатуры незначительна. Однако Са2+ играет важную роль в возникновении ПД сердечной и гладкой мышц, в передаче импульсов от одного нейрона к другому, от нервного волокна к мышечному, в обеспечении мышечного сокращения.
4. Следовая гиперполяризация клеточной мембраны (рис. 1.3 -4) обычно является следствием еще сохраняющейся повышенной проницаемости клеточной мембраны для К+, она характерна для нейронов. Ворота К-каналов еще не полностью закрыты, поэтому К+ продолжает выходить из клетки согласно концентрационному градиенту, что и ведет к гиперполяризации клеточной мембраны. Постепенно проницаемость клеточной мембраны возвращается к исходной (натриевые и калиевые ворота возвращаются в исходное состояние), а мембранный потенциал становится таким же, каким он был до возбуждения клетки. Ыа/ К-помпа непосредственно за фазы потенциала действия не отвечает, хотя она и продолжает работать во время развития ПД.
Следовая деполяризация также характерна для нейронов, она может быть зарегистрирована и в клетках скелетной мышцы. Механизм ее изучен недостаточно. Возможно, это связано с кратковременным повышением проницаемости клеточной мембраны для Ыа+ и входом его в клетку согласно концентрационному и электрическому градиентам.
В. Запас ионов в клетке, обеспечивающих возникновение возбуждения (ПД), огромен. Концентрационные градиенты ионов в результате одного цикла возбуждения практически не изменяются. Клетка может возбуждаться до 5105 раз без подзарядки, то есть без работы Ыа/К-насоса. Число импульсов, которое генерирует и проводит нервное волокно, зависит от его толщины, что определяет запас ионов. Чем толще нервное волокно, тем больше запас ионов и больше импульсов оно может генерировать (от нескольких сот до нескольких сотен тысяч) без участия №/К-насо-са. Однако в тонких С-волокнах на возникновение одного ПД расходуется около 1 % концентрационных градиентов №+и К+. Таким образом, если заблокировать выработку энергии, то клетка будет еще многократно возбуждаться и в этом случае. В реальной же действительности Ыа/К-насос постоянно переносит Ыа+ из клетки, а К+ возвращает в клетку, в результате постоянно поддерживается концентрационный градиент №+ и К+, что осуществляется за счет непосредственного расхода энергии, источником которой является АТФ.
ЛОКАЛЬНЫЙ ПОТЕНЦИАЛ
При раздражении возбудимой ткани не всегда возникает ПД. В частности, если сила раздражителя мала, деполяризация не достигнет критического уровня, естественно, не возникнет импульсное, распространяющееся возбуждение. В этом случае ответ ткани на раздражение носит форму локального потенциала. Локальными потенциалами возбудимых клеток также являются: возбуждающий постсинаптический потенциал, рецепторные потенциалы, тормозной постсинаптический потенциал. Величина локальных потенциалов весьма вариабельна, она может достигать 10-40 мВ в зависимости от рода клеток и силы стимула. Свойства такого ответа существенно отличаются от импульсного (табл. 1.1).

|
Повышение возбудимости клетки во время локального потенциала объясняется тем, что клеточная мембрана оказывается частично деполяризованной. Если Е остается на постоянном уровне, для достижения критического уровня деполяризации во время локального потенциала нужен значительно меньшей силы раздражитель. Амплитуда ПД не зависит от силы раздражения, потому что он возникает вследствие регенеративного процесса. (О причине невозбудимости клетки при возникновении ПД см. в разделе 3.4.)

