




В 1939 г. американские[Мф54] физиологи Джозеф Эрлангер [Б55] и Герберт С.Гассер [Б56] [Б57] зарегистрировали токи действия от целого нервного ствола седалищного нерва лягушки на разных расстояниях от стимулирующего электрода (рис. 210041905).[Б58]

Было установлено, что регистрируемый суммарный потенциал имеет ряд пиков, которые были обозначены буквами латинского алфавита A, B, C (рис., I). Пик A имел дополнительные пики, помеченные греческими буквами α, β, γ, δ (рис., II). [Б59] В 1944 г. работа Дж.Эрлангера и Г.С.Гассера была оценена присуждением Нобелевской премии[Б60].

Нервы у позвоночных состоят из трех основных групп волокон (А, В и С), различающихся по степени миелизации, диаметру волокна, длительности пика ПД (скорости развития ПД), электровозбудимости, его компенсации и скорости проведения (все эти показатели в ряду А — В — С падают).
Группа А включает наиболее толстые хорошо миелинизированные моторные и чувствительные волокна; группа В — слабомиелинизированные, преганглионарные волокна автономной нервной системы; группа С - немиелинизированные, постганглионарные (симпатические) волокна.
В группе А в ряду a, b, g, d названные показатели тоже падают. Соотношения свойств этих групп волокон демонстрируются в табл..
Необходимо заметить, что указанные соотношения порогов электрического раздражения групп волокон не отражают точного соотношения электровозбудимости их мембран. Относительно высокие пороги тонких волокон при их раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие волокна по сравнению с толстыми обладают более высоким входным сопротивлением. В них входит такая малая часть раздражающего тока, что при пороговой силе для Аa -волокон она совершенно недостаточна для создания на мембране более тонких волокон сколько-нибудь существенной деполяризации. По этой же причине (высокое RI) отводимые от ствола (внеклеточно) ПД тонких волокон предстают значительно меньшими, чем ПД толстых волокон.

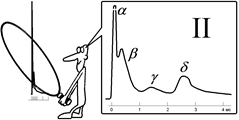
Рис.. Составные части потенциала действия смешанного нерва.
I – при относительно медленной скорости записи.
II – при относительно высокой скорости записи.
Объяснение в тексте. По оси абсцисс – время, по оси ординат амплитуда составного потенциала в мВ.
Суммарная электрическая активность нерва создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и временным параметрам ПД, распространяющийся в обе стороны от точки, к которой приложено раздражение. Суммарный электрический сигнал нерва зависит от числа активных волокон, синхронности их активности, способа отведения и других обстоятельств.
Рассмотрим случай так называемого однофазного отведения, при котором один отводящий электрод (активный) расположен на нормальном участке ствола, а другой (индифферентный) — на поврежденном, где волокна деполяризованы полностью (рис.1.19). Пусть применяют искусственное раздражение и раздражающий электрод (катод) находится достаточно близко (l= 3 мм) от активного отводящего электрода, а нерв помещен в непроводящую среду (масло или воздух). Здесь сильное одиночное раздражение приводит к синхронному возбуждению всех волокон, при этом активный отводящий электрод регистрирует суммарный ПД нерва, по форме приближающийся к ПД отдельного A(альфа)‑волокна, но немного более затянутый во времени. Этот ПД нерва, однако, не подчиняется правилу «все или ничего». При пороговом раздражении он ничтожно мал, с увеличением силы стимулов постепенно растет, достигая максимума, равного при обычных условиях 5—10 мВ, а в условиях сахарозного мостика — 50—100 мВ.
При дальнейшем увеличении силы стимула этот ПД несколько удлиняется во времени. Все изменения амплитуды и длительности пика ПД нерва при усилении стимула определяются ростом числа активных волокон, подключением к низкопороговым и быстрым А(альфа)-волокнам более высокопороговых медленных бета-, гамма-, дельта-волокон группы А, затем В- и, наконец, С-группы.
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Скорость проведения, м/с |
| Aa | 13 - 22 | 70 - 120 |
| Ab | 8 -13 | 40 - 70 |
| Ag | 4 - 8 | 15 – 40 |
| Ad | 1 – 4 | 5 – 15 |
| B | 1 - 3 | 3 – 14 |
| C | 0,5 – 1,0 | 0,5 - 2 |
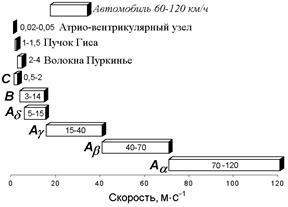
Таблица. Классификация нервных волокон по Дж.Эрлангеру и Х.Гассеру
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Пороги электрического раздраженния (относительно Aa) | Длительность пика ПД1 | Отрицательный следовой потенциал (ОСП) | Положительный следовой потенциал [Б61] | Скорость проведения, м/с | ||
| Длительность, мс | Амплитуда СП, % к амплитуде ПД | Длительность, мс | Амплитуда СП, % к амплитуде ПД | |||||
| Aa | 13 - 22 | 1,0 | 0,4 | 15 – 20 | 40 - 60 | 0,2 | 70 - 120 | |
| Ab | 8 -13 | 40 - 70 | ||||||
| Ag | 4 - 8 | 15 – 40 | ||||||
| Ad | 1 – 4 | 5 – 15 | ||||||
| B | 1 - 3 | 11,7 | 1,2 | ОСП нет | 100 – 300 | 3 – 14 | ||
| C | 0,5 – 1,0 | 100,0 | 2,0 | 50 – 60 | 300 - 1000 | 0,5 - 2 |
1Приблизительно ту же величину имеют и абсолютные рефрактерные фазы

