




Большое преимущество при сравнительном изучении поведения дает то, что нам уже известна в общих чертах естественная система филогенетического родства, т. е. здесь наше положение гораздо предпочтительнее, чем у морфологов лет триста назад. Если обнаружится, например, что социальная организация, лежащая в основе брачного поведения у каракатицы, очень сходна с известной для рыб, мы ни на секунду не признаем это доказательством родства между ними, поскольку морфологические исследования уже давно позволили отнести рыб и каракатиц к совершенно различным типам животных. Некоторые черты их морфологического сходства типа наличия «плавников» или глаз — конвергентные признаки, как и особенности брачного поведения.
С другой стороны, сравнивая брачное поведение близкородственных видов, вполне можно предположить, что его схемы у них гомологичны. Так, в ходе зигзагообразного танца самец трехиглой колюшки сначала ведет самку, а затем нападает на нее, в случае десятииглой колюшки — сначала атакует, затем ведет, а у пятнадцатииглой колюшки (Spinachia vulgaris) — просто атакует и никуда не ведет, пока она сама не проявит инициативу. Сразу же напрашивается вывод о трех формах единого в своей основе поведения. [117] Если же брачные церемонии близкородственных видов сильно различаются, можно попытаться обнаружить общий предковый тип, от которого они эволюционировали.
Поведение редко систематически изучали с этой точки зрения. Однако именно социальные взаимоотношения дают исследователю уникальную возможность проследить ход эволюции, поскольку в связи с потребностью в репродуктивной изоляции тенденция к разнообразию здесь обычно проявляется в первую очередь. Другими словами, социальная организация родственных видов дивергировала очень быстро, а значит, даже близкие формы (которые легче всего сравнивать, поскольку гомологии очевидны) должны заметно различаться своим поведением.
Как и в случае морфологии, сравнение можно проводить на любом уровне: сообщества в целом, брачного поведения или отдельного релизера. В любом случае должно хватить данных для того, чтобы прийти к четким эволюционным выводам.
Сравнивая социальные организации различных пчел, нетрудно обратить внимание на преобладание одиночных видов и исключительность медоносной пчелы (и двух близкородственных ей таксонов), живущей в сложно устроенных «государства», основанных на сотрудничестве тысяч особей. Следовательно, логично допустить, что исходно пчелы были одиночными. Как указывалось в гл. 7, у некоторых родственных пчелам насекомых существует определенный уровень социальности, т.е. они занимают промежуточное в этом смысле положение. Сравнивая одиночные, средне- и высокосоциальные таксоны, можно видеть, что социальная организация развивалась из семьи, включающей самку и ее потомство, причем сначала возникла ассоциация между ними, а затем появилось разделение труда особей с постепенным усложнением сотрудничества.
Исследовать этот вопрос на муравьях труднее, потому что среди них нет одиночных видов, как и у термитов. Сравнение муравьев и термитов четко демонстрирует конвергенцию между этими отдаленными группами. Социальная организация термитов возникла иным путем, чем у муравьев или пчел, поскольку у этих «общественных тараканов» во всех кастах представлены самцы, а государство возникло из семьи, включающей самца, самку и их потомство. Хорошо известно, что параллели между муравьями и термитами можно провести по многим признакам; например, и у тех и у других выделяется каста «солдат».
Переходя теперь от социальной организации в целом к ее деталям, можно опять же видеть гомологии и конвергенцию. Они особенно бросаются в глаза в случае брачного поведения. У групп с хорошо развитым зрением часто наблюдается половой диморфизм: самцы броско окрашены или выполняют особые демонстрационные церемонии. Это свойственно, например, манящим [118] крабам (рис. 66), колюшкам, бойцовым рыбкам

Рис. 64. Рабочий муравей (слева) и солдат.
Рис. 65. Рабочий термит (слева) и солдат.
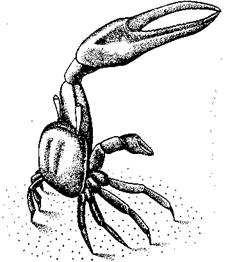
Рис. 66. Самец манящего краба во время демонстрации.
(Betta splendens), ящерицам и многим птицам. Все они используют свою яркую окраску, чтобы угрожать другим самцам, причем их потенциально противоречивое поведение (либо драка, либо ухаживание за другими особями своего вида) направляется в чисто половое русло особыми реакциями самки. Некоторые самцы преимущественно агрессивны (например, бойцовые рыбки или голуби), у других преобладают половые тенденции, и их провоцируют на драку специальные релизеры соперников. Это свойственно, например, каракатице и мускусной утке. У некоторых птиц существует [119] любопытная «токовая» система, при которой множество ярко окрашенных самцов собираются на общую площадку для брачных демонстраций (ток), посещаемую самками только для совокупления. Она развивалась независимо у турухтана (Philomachus pugnax) и тетерева (Lyrurus tetrix). У обоих этих видов самцы не присоединяются к самкам, чтобы помочь им в выполнении родительских обязанностей; «личных» связей у этих птиц не возникает вообще.
Внутри одного рода часто можно проследить гомологии. На первый взгляд поведение при образовании пар у серебристой и обыкновенной чаек выглядит совершенно различно. В первом случае партнеры находят друг друга в «клубах», т. е. в местах скопления многих особей, во втором — на так называемых «предтерриториях». Одинокий самец обыкновенной чайки очень агрессивно реагирует на любого сородича независимо от его пола, а у серебристой чайки он, нападая на других самцов, вполне терпим к самкам. Обыкновенная чайка использует воздушную демонстрацию, серебристая — нет. Только что образовавшаяся пара обыкновенной чайки улетает с места «знакомства» выбирать гнездовой участок, а серебристые чайки удаляются от клуба пешком и часто вьют гнездо недалеко от него. В некоторых деталях поведения также заметны существенные различия; угрожающая поза неодинакова: у серебристой чайки она «вертикальная», а у обыкновенной — «впередсмотрящая». Различны и умиротворяющие действия: у серебристой чайки — особая «поза подчинения», а у обыкновенной — «отмашка головой».
Однако более пристальный анализ способа образования пар у обоих видов говори!, что общая схема здесь одинакова: самки приближаются к самцам и умиротворяют их, принимая позу, противоположную угрожающей, после чего партнеры вместе выбирают гнездовую территорию. Различия связаны с двумя обстоятельствами: во-первых, обыкновенная чайка как меньшая по размеру больше времени проводит в полете, поэтому у нее сформировались воздушные демонстрации, отсутствующие у более крупной серебристой чайки, иная угрожающая поза («вертикальная» ориентирована на противника, находящегося на земле, а «вперед смотрящая» — на чужака, который может и подойти, и подлететь), она по-особому отправляется на поиски гнездового участка. Во-вторых, угрожающая поза у обыкновенной чайки подкрепляется коричневой окраской ее «лица», которая обусловливает и «отмашку головой» в качестве умиротворяющей церемонии.
Наши знания о таких вещах еще очень фрагментарны и совершенно недостаточны для реконструкции эволюционных процессов, приведших к появлению различных типов поведения. [120]
Сравнение релизеров.
Несколько больше известно об отдельных сигналах, регулирующих поведение. Здесь опять же нетрудно обнаружить гомологии и конвергенцию. «Смещенная» чистка перьев у ухаживающего за самкой самца утки, хотя и неодинакова у разных видов, остается явно «одним и тем же» во всех случаях. Пение птиц, специфическое у каждого вида из-за необходимости в репродуктивной изоляции, гомологично у всей группы певчих, как и используемый для этого орган — нижняя гортань. Примеры конвергенции — это, в частности, фронтальная демонстрация у рыб, поднимающих жаберные крышки, и раздувание в ярко окрашенный веер или воротник шейных перьев у самцов турухтана и домашнего петуха.
Сравнение гомологичных релизеров привело к интересным выводам об их происхождении и эволюции.
До сих пор выделено два источника сигнальных движений. Один из них — это движение намерения, или интенционное. Когда утка или гусь собираются взлететь, их мотивация формируется постепенно. Когда она очень слабая, можно видеть интенционные движения. Оперение прижимается к телу, после чего голова неоднократно подергивается; это — самая низкая по интенсивности готовность к взлету. При усилении мотивации подергивания головой усиливаются и в движение могут прийти другие части тела: крылья приподнимаются, а тело иногда немного наклоняется вперед. Такие слабовыраженные формы интенционного движения действуют на сородичей как релизеры.
В других случаях животное может совершать интенционные движения даже при довольно сильной мотивации. Вертикальная угрожающая поза серебристой чайки, несомненно, указывает на достаточно выраженную тенденцию к нападению, однако перерасти в настоящую атаку ей нелегко, так как этому противодействует одновременная тенденция к бегству или отступлению. Такие подавленные интенционные движения действуют как сигналы и во многих других случаях.
Второй источник релизеров — смещенная активность. Ее примеры — дерганье травы серебристой чайкой, угрожающее рытье песка трехиглой колюшкой или указывание ею же входа в гнездо (смещенное «обмахивание» икры). Все эти действия играют роль сигналов, запускающих определенные реакции у противника или полового партнера.
Каким образом другие особи начали «понимать» такие движения, проследить трудно. Эта проблема связана с происхождением реактивности на сигнал, а не самого сигнального движения. Что касается интенционных движений, вопрос ставится в сущности о происхождении реакции животного на любой внешний стимул. Почему черный дрозд реагирует на земляного червя или на [121] ястреба-перепелятника — не меньшая загадка, чем причина его реакции на интенционное движение другого черного дрозда, служащее предупреждением об опасности.
Почему серебристая чайка «понимает» агрессивную природу смещенного сбора гнездового материала (дерганья травы) и не реагирует на него, сама занимаясь строительством гнезда,— вопрос иного порядка. По-моему, причин для интерпретации такого поведения как агрессивного две. Во-первых, оно чередуется с настоящими агрессивными действиями. Во-вторых, как уже говорилось, это дерганье отличается от обычного сбора гнездового материала: чайка клюет траву «яростно» и тянет ее изо всех сил. Такие детали характерны для драки; птица угрожает растениям, как будто это ее противник.
Когда сформировалась реакция на сигнальное движение другого индивида, дальнейшее развитие его знаковой функции зависит как от агента, так и от реагента. У обоих животных начинается новый адаптивный эволюционный процесс. Сравнение позволяет выявить несколько его аспектов. Смещенное охорашивание, т. е. сигнальное движение, составляющее часть ухаживания у многих самцов уток, у каждого из их видов несколько отличается от обычной чистки перьев, причем в некоторых случаях разница настолько велика, что природу смещенного движения понять трудно. Лоренц привел детальные описания и иллюстрации, основанные на киносъемке смещенного охорашивания у многих видов (рис. 67). Относительно примитивна его форма у кряквы. Самец просто прячет клюв под крыло почти так же, как при обычной чистке перьев, но смещенное движение более стереотипно. У мандаринки оно весьма специализировано: самец ловко дотрагивается до опахала одного из маховых перьев второго порядка, которое отличается от обычных перьев темно-зеленого цвета. Его опахало преобразовано в огромную флагоподобную структуру, а окраска ярко-оранжевая. У чирка-трескунка это движение опять же иное. Он дотрагивается не до внутренней, а до наружной стороны крыла как раз в том месте, где на нем находятся яркие серовато-голубые кроющие перья. Таким образом, и у него, и у мандаринки развилась бросающаяся в глаза структура, и движение привлекает внимание именно к ней. Эволюция привела к тому, что оно стало более заметным и стереотипным, превратилось, так сказать, в «ритуал». Одновременно произошла дивергенция этих движений у разных таксонов, повысилась их видоспецифичность. Эволюционный процесс, «ухватывающий» сигналы и делающий их более заметными и видоспецифичными, называется ритуализацией.
Все накопленные до сих пор данные приводят к выводу, что сигнальные движения первоначально не обладали знаковой функцией. В каком-то смысле это были «побочные продукты» нервной [122] организации. После появления знаковой функции начался новый тип адаптивной эволюции, ритуализация, приведшая к одновременному изменению самого движения и связанной с ним морфологической структуры.

Рис. 67. Смещенная чистка перьев ухаживающими утиными: 1 — пеганка; 2 — чирок-трескунок; 3 — мандаринка; 4— кряква.
Ритуализация адаптивна с двух точек зрения. Ритуализованные релизеры всегда хорошо заметны и просты. Это — адаптация к ограниченной реактивности врожденного поведения. Каждая врожденная реакция требует для своего запуска специфических стимулов; их изучение показало, что они всегда относительно просты и броски. Ритуализация обычно ведет к специализации релизера на предъявлении таких «знаковых стимулов», т. е. релизеры в определенном смысле их «материализуют». Во-вторых, ритуализация направлена на то, чтобы каждый данный релизер отличался от любого другого как у этого, так и у другого таксона. В результате облегчается социальное сотрудничество сородичей и снижается вероятность реакции на другой вид. [123]
В случае интенционных движений и смещенной активности ритуализация, по-видимому, развивается одинаково. В обоих случаях главный упор может делаться как на движение, так и на структуру. Одно из самых обычных изменений движения — его «схематизация», подразумевающая подчеркивание одних компонентов и ослабление других. Это можно наблюдать, например, на некоторых ухаживающих действиях у уток. Так, «укорачивание» тела первоначально означало подъем головы и хвоста. У чирка-трескунка движение головы назад усилилось, а хвоста вверх — полностью исчезло. У чилийского чирка главный акцент сделан на движение груди, а хвост опять же не поднимается. У шилохвости задействованы и голова, и хвост; движение последнего подчеркивается ярко окрашенным треугольником у его основания и вытянутой формой.
Физиологически эти и другие аспекты ритуализации можно истолковать как количественные изменения с пороговыми значениями для различных составных частей движения. Более глубокое обсуждение этих проблем увело бы нас за рамки выбранной тематики. Однако мне хотелось бы отметить, что изучение эволюции релизеров очень важно для понимания происхождения и развития «новых» элементов поведения в целом, поскольку ритуализация интенционных движений или смещенных действий ведет как раз к этому. Углубляться в данную проблему мы также не будем.
Заключение.
Изучение происхождения и эволюции релизеров, в частности используемых при ухаживании и угрозе, не оставляет сомнений в том, что первоначально они были случайными «побочными продуктами», разрядкой нервного возбуждения в форме интенционных движений или смещенной активности. В большинстве случаев такие формы разрядки возникают, когда нормальные ее способы подавляются одновременной активацией противоположных по направленности побуждений. Это справедливо для всех «подавленных интенционных движений», всех угрожающих смещенных действий и, возможно, для многих форм смещенной активности при ухаживании. По-видимому, такой вывод позволяет понять, почему «демонстрации» так широко распространены именно при ухаживании и угрозе. В первом случае, если половое влечение и остается главным компонентом мотивации, все же одновременно проявляются агрессивность и тенденция к избеганию. При угрозе в конфликт между собой вступают два последних побуждения. Уже говорилось, что как агрессивность, так и половое влечение необходимы для сохранения вида. Поскольку врожденное поведение складывается из реакций на простые знаковые стимулы, а самка, будучи представителем того же вида, что и самец, [124] помимо стимулов, вызывающих у него половую реакцию, не может не посылать сигналов, провоцирующих агрессивность, при приближении ее к партнеру у того всегда активируются и враждебность, и сексуальность. Если бы агрессивность была слабее, он мог бы реагировать на самку только как на полового партнера, но за счет снижения эффективности драк с другими самцами. Если усилить половое влечение, оно бы преодолело не только враждебность к самке, но и прочие побуждения, включая страх перед хищником. Если бы слабее проявлялась тенденция к избеганию, это оказалось бы полезным при внутривидовых столкновениях, но повысило бы уязвимость при встрече с хищником. Таким образом, в каждом животном одни побуждения точно уравновешивают другие. Угрожающее поведение и ухаживание — неизбежные последствия такого равновесия; ритуализация делает их максимально приспособленными к условиям существования вида. [125]

