




Аеробне й анаеробне перетворення вуглеводів тісно зв'язані між собою. Це насамперед виявляється в тому, що обидва процеси проходять однаково включно до стадії утворення піровиноградної кислоти. В них беруть участь одні й ті самі ферменти та утворюються однакові проміжні продукти. Відмінність між анаеробним і аеробним розкладанням вуглеводів починається з перетворення піровиноградної кислоти.
Якщо вуглеводи перетворюються в анаеробних умовах, то піровиноградна кислота, як уже зазначалося, відновлюється до молочної кислоти. При перетворенні вуглеводів в аеробних умовах піровиноградна кислота піддається декарбоксилюванню з утворенням ацетил-KoA, який далі окислюється до кінцевих продуктів – CO2 і H2O з виділенням значної кількості енергії, що акумулюється в молекулах ATФ.
Процес перетворення піровиноградної кислоти до ацетил-КоА дістав назву окислювального декарбоксилування. Каталізується він складним поліферментним комплексом – піруватдегідрогеназою, який пов'язаний з дією п'яти коферментів: тіамінпірофосфату, ліпоєвої кислоти, коензиму А, НАД і ФАД. Дослідженнями доведено, що окислювальне декарбоксилування піровиноградної кислоти в тканинах організмів людини і тварин проходить лише в аеробних умовах. Загальну схему можна подати так:

У зв'язку зі значним зменшенням вільної енергії ця реакція є необоротною. Вона проходить у декілька стадій.
На першій стадії піровиноградна кислота взаємодіє з тіамінпірофосфатом з утворенням проміжної сполуки, де вона перебуває в активному стані. Далі під каталітичною дією піруватдекарбоксилази піровиноградна кислота, що входить до складу тіамінпірофосфату, декарбоксилується. В результаті цієї реакції утворюється оксіетилтіамінпірофосфат, який під впливом ферменту ліпоїлредуктази ацетил-трансферази, коферментом якого є ліпоєва кислота, розкладається на вільний тіамінпірофосфат і ацетилліпоєву кислоту. Ацетилліпоєва кислота утворюється внаслідок переходу залишку оцтової кислоти від оксіетилтіамінпірофосфату до ліпоєвої кислоти:

З ацетилліпоєвої кислоти залишок ацетилу далі переноситься на KoA–SH. Продуктами реакції є ацетил-КоА і дигідроліпоєва кислота. Слід підкреслити, що дигідроліпоєва кислота під впливом ФАД здатна переходити у ліпоєву кислоту:
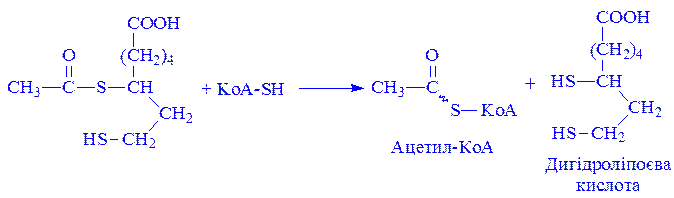

За цих умов відновлений ФАД передає атоми водню на окислений НАД:

Відновлений НАД, що утворився, за допомогою цитохромної системи окислюється киснем повітря:
НАД×H2 + [O] ® H2O + НАД+.
У результаті окислення, спряженого з процесами фосфорилування, синтезується три молекули АТФ, тобто акумулюється 126 кДж енергії.
Частина ацетил-КоА, що утворився під час декарбоксилювання піровиноградної кислоти, використовується для синтезу жирів, вуглеводів та інших сполук, а інша вступає в цикл трикарбонових кислот, де окислюється до CO2 і H2O. При цьому вивільнюється певна кількість енергії.
Цикл трикарбонових кислот Кребса (ЦТК)
Основна маса хімічної енергії вуглеводів звільняється в аеробних умовах за участю кисню. Цикл трикарбонових кислот Кребса називають ще циклом лимонної кислоти, або клітинним диханням.
Між анаеробним і аеробним розщепленням вуглеводів існує тісний зв'язок. Перш за все це виявляється в наявності загальної зв'язуючої ланки – піровиноградної кислоти, якою завершується анаеробне розщеплення вуглеводів і починається клітинне дихання. Обидві фази каталізують одні і ті ж ферменти (дегідрогенази, кінази, ін.). Хімічна енергія, яка вивільняється при фосфорилуванні, резервується у вигляді макроергів АТФ. У хімічних реакціях беруть участь одні і ті ж коферменти (НАД, НАДФ) і катіони. Відмінності полягають в наступному. Якщо ферменти анаеробного розщеплення вуглеводів локалізуються в гіалоплазмі, то реакції клітинного дихання протікають переважно в мітохондріях – енергетичних підстанціях клітини. За певних умов виявляється антагонізм між обома фазами. Так, у присутності кисню швидкість реакцій гліколіза різко зменшується (ефект Пастера). Продукти гліколіза можуть гальмувати аеробний обмін вуглеводів (ефект Кребтрі).
Цикл трикарбонових кислот є ланцюгом послідовних хімічних реакцій, у результаті яких продукти розщеплення вуглеводів окислюються до CO2 і Н2О, а хімічна енергія акумулюється у вигляді макроергічних сполук. У ході клітинного дихання утворюється „носій” – щавелевооцтова кислота (ЩОК). Далі відбувається конденсація з „носієм” активованого залишку оцтової кислоти. Утворюється трикарбонова кислота – лимонна. В ході хімічних реакцій відбувається кругообіг залишку оцтової кислоти в циклі. Під час функціонування циклу мають місце процеси дегідрування і декарбоксилування. З кожної молекули піровиноградної кислоти утворюється 18 молекул АТФ. В кінці циклу звільняється „носій”, який вступає в реакцію з новими молекулами активованого залишку оцтової кислоти. Розглянемо послідовність реакцій ЦТК:
1. Якщо кінцевим продуктом анаеробного розщеплення вуглеводів є молочна кислота, то під впливом лактатдегідрогенази вона окислюється в піровиноградну кислоту:

2. Частина молекул піровиноградної кислоти йде на синтез „носія” ЩОК під впливом ферменту піруваткарбоксилази і присутності іонів Mg2+:

3. Інша частина молекул піровиноградної кислоти служить джерелом утворення „активного ацетату” (окислювальне декарбоксилування) – ацетилкоензиму А (ацетил–KoA). Реакція протікає під впливом піруватдегідрогенази:
Ацетил-КоА має в своїй молекулі макроергічний зв'язок, в якому акумулюється близько 5 –7% енергії. Основна маса хімічної енергії утворюється в результаті окислення „активного ацетату”.
4. Під впливом цитратсинтази починає функціонувати власне цикл трикарбонових кислот, що призводить до утворення лимонної кислоти:

5. Лимонна кислота під впливом ферменту аконітатгідратази дегідрується і перетворюється на цис -аконітову кислоту, яка після приєднання молекули води перетворюється в ізолимонну:

Між трьома трикарбоновими кислотами встановлюється динамічна рівновага. Наступні реакції протікають за рахунок ізолимонної кислоти, на яку перетворюються дві інші.
6. Ізолимонна кислота окислюється в щавелевоянтарну кислоту, яка декарбоксилується і перетворюється в a-кетоглутарову кислоту. Реакцію каталізує фермент ізоцитратдегідрогеназа:

7. a-Кетоглутарова кислота під впливом ферменту 2-оксо-(a-кето)-глутаратдегідрогенази декарбоксилується, внаслідок чого утворюється сукциніл-КоА, що містить макроергічний зв'язок:

8. На наступній стадії сукциніл-КоА під впливом ферменту сукциніл-КоА – синтетази передає макроергічний зв'язок ГДФ (гуанозиндифосфату):

ГТФ під впливом ферменту ГТФ-аденілаткінази віддає макроергічний зв'язок АМФ:
ГТФ + АМФ ® ТДФ + АДФ.
9. Янтарна кислота під впливом ферменту сукцинатдегідрогенази (СДГ) окислюється до фумарової. Коферментом СДГ є ФАД:

10. Фумарова кислота під впливом ферменту фумаратгідратази перетворюється на яблучну:

11. Яблучна кислота під впливом ферменту малатдегідрогенази (МДГ) окислюється, утворюючи ЩОК:

За наявності в реагуючій системі ацетил-КоА ЩОК знову включається в цикл трикарбонових кислот (див. 4-у стадію).
Таким чином, відносно невелика кількість щавелевооцтової кислоти, вступаючи кілька разів у реакцію конденсації з ацетил-KoА, забезпечує окислення значної кількості оцтової кислоти, яка утворюється в процесі обміну не тільки вуглеводів, а й ліпідів і білків. Внаслідок усіх перетворень циклу Кребса оцтова кислота у вигляді ацетил-КоА розкладається на CO2 і H2O:
CH3–COOH + 2O2 = 2CO2 + 2H2O.
Енергія, що вивільнюється під час окислення оцтової кислоти, акумулюється в макроергічних зв'язках АТФ (схема 4).

Схема 4. Цикл трикарбонових кислот Кребса (Q – місця гальмування циклу фторацетатом, малонатом, арсенатом)
Як видно з рівнянь реакцій, у циклі Кребса утворюється чотири молекули відновлених форм коферментів: одна молекула НАДФ×H2, дві молекули НАД×H2 і одна молекула ФАД×H2. При цьому доведено, що під час окислення однієї молекули НАД×H2 або НАДФ×H2 шляхом відщеплення атома водню в ланцюгу дихальних ферментів синтезується три молекули АТФ. Внаслідок окислення ФАД×H2 утворюється дві молекули АТФ. Отже, всього під час окислення відновлених форм коферментів утворюється 3 ´ 3 + 2 = 11 молекул АТФ. Крім того, одна молекула АТФ утворюється на рівні субстрату під час перетворення сукциніл-КоА в янтарну кислоту.
У процесі перетворення однієї молекули ацетил-КоА в циклі трикарбонових кислот синтезується 11 + 1 = 12 молекул АТФ. Разом з тим три молекули АТФ синтезуються внаслідок окислення однієї молекули НАД×H2, яка утворюється в процесі перетворення піровиноградної кислоти до ацетил-KoА. Всього в процесі перетворення однієї молекули піровиноградної кислоти до ацетил-KoА і останнього до CO2 і H2O синтезується 12 + 3 = 15 молекул АТФ. Оскільки з однієї молекули глюкози утворюється дві молекули піровиноградної кислоти, то всього утворюється 15 ´ 2 = 30 молекул АТФ. Крім того, 6 молекул АТФ утворюється внаслідок окислення двох молекул НАД×H2, які вивільнюються під час гліколітичної оксиредукції, і дві молекули синтезується в процесі гліколізу. Toму загальний енергетичний ефект аеробного розкладання однієї молекули глюкози до кінцевих продуктів – CO2 і Н2О – становить 30 + 6 + 2 = 38 молекул АТФ. Оскільки в одній молекулі АТФ зосереджено 42 кДж енергії, то всього під час аеробного перетворення однієї молекули глюкози акумулюється 1596 кДж енергії. Перетворення однієї молекули глюкози за анаеробних умов, як зазнавалось раніше, дає лише дві молекули ATФ, тобто в макроергічннх зв'язках її акумульовано 2 ´ 42 = 84 кДж енергії.
Отже, основним джерелом енергії для організму є аеробне окислення органічних сполук. Коефіцієнт корисної дії ЦТК рівний 0,5. Решта енергії розсіюється у вигляді теплоти. У ЦТК в середньому окислюється 16 – 33% молочної кислоти, інша частина використовується для ресинтезу глікогену.
Пентозний цикл
Пентозний цикл – це ланцюг послідовних хімічних перетворень вуглеводів, у результаті якого в тканинах і клітинах звільняється хімічна енергія і утворюються пентози, необхідні для синтезу нуклеїнових кислот, нуклеотидів і коферментів. Його часто називають апотомічним циклом, оскільки при окисленні глюкози відщеплюється один атом вуглецю. Іноді його називають прямим, або гексозомонофосфатним шляхом окислення вуглеводів, оскільки тут глюкозо-6-фосфат піддається прямому окисленню (з відщепленням CO2) без утворення фруктозо-1,6-дифосфата і двох фосфотріоз (схема 5).

Схема 5. Перетворення пентозофосфатів
Частка пентозного циклу в кількісному перетворенні глюкози невелика. У ссавців активність циклу висока в ембріональних тканинах, печінці, наднирниках, пухлинах, молочній і щитовидній залозах, еритроцитах. Пентозний цикл дає організму відновлений НАДФ – донор водню для біосинтезу жирних кислот, холестерину, пуринових і піримідинових основ, кетостероїдів, глюкози, ін. Пентозний цикл складається з таких стадій:
1. Глюкозо-6-фосфат, що утворюється в результаті фосфороліза глікогену або фосфорилування глюкози, під впливом глюкозо-6-фосфатдегідрогенази окислюється і перетворюється в 6-монофосфоглюкон:

2. 6-Монофосфоглюкон під впливом ферменту глюконолактонази приєднує молекулу води, перетворюючись на 6-фосфоглюконову кислоту:
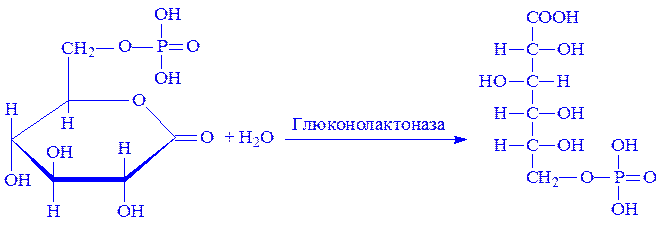
3. 6-Фосфоглюконова кислота за участю фосфоглюконатдегідрогенази піддається окислювальному декарбоксилуванню, що призводить до утворення кетопентози – D-рибулозо-5-фосфата і другої молекули НАДФ×H2:

4. D-Рибулозо-5-фосфат під впливом ферменту рибулозофосфат-3-епімерази оборотно перетворюється на свій епімер – D-ксилулозо-5-фосфат (а). У деяких випадках D-рибулозо-5-фосфат може оборотно перетворюватися на свій альдоізомер – D-рибозо-5-фосфат (б):
| а) | 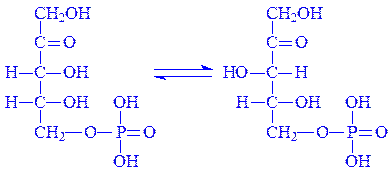
|
| б) | 
|
D-Рибозо-5-фосфат використовується клітинами для синтезу РНК і нуклеотидів (АМФ, АДФ, АТФ). Часто пентозний цикл на даній стадії завершується. Його можна підсумувати рівнянням:
Глюкозо-6-фосфат + 2НАДФ+ Û D-Рибозо-5-фосфат + CO2 +2HАДФ×H2
Пентози, які не використані для синтезу нуклеїнових кислот і нуклеотидів, витрачаються на біосинтез інших сполук, регенерацію глюкози, з якої починався пентозний цикл.
5. З D-кcилулозо-5-фосфата і D-рибозо-5-фосфата під впливом транскетолази може утворюватися D-седогептулозо-7-фосфат і 3-фосфогліцериновий альдегід:
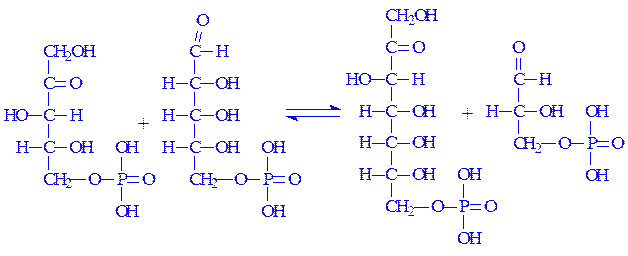
6. 3-
Фосфогліцериновий альдегід може включатися в 4-у стадію анаеробного розщеплення вуглеводів або під впливом ферменту трансальдолази взаємодіяти з D-седогептулозо-7-фосфатом, утворюючи фруктозо-6-фосфат і еритрозо-4-фосфат:

Фруктозо-6-фосфат може ізомеризуватися в глюкозо-6-фосфат і вступати в 1-у стадію пентозного циклу або включатися в 2-у стадію анаеробного розщеплення вуглеводів.
7. Еритрозо-4-фосфат під впливом ферменту транскетолази взаємодіє з ксилулозо-5-фосфатом, що призводить до утворення фруктозо-6-фосфата і 3-фосфогліцеринового альдегіду (їх перетворення див. вище):

Реакції пентозного циклу можна підсумувати рівнянням:
6 Глюкозо-6-фосфат + 7Н2О + 12НАДФ+ ® 6СО2 + 12НАДФ×Н2 + 5 Глюкозо-6-фосфат + H3PO4.
Отже, виходячи із сумарного рівняння бачимо, що при повному окисленні 1 молекули глюкози утворюється 12 молекул відновленого НАДФ, які в процесі окислення у мітохондріях можуть дати 12 ´ 3 = 36 молекул АТФ. У макроергічних зв’язках 36 молекул АТФ зосереджено 36 ´ 42 = 1512 кДж енергії. Однак останнім часом прийнято вважати, що утворені за пентозним циклом відновлені форми НАДФ використовуються в основному в процесах синтезу різних сполук (жирних кислот, холестерину, тощо) в організмі.
Пентозному циклу належить важлива роль в синтезі жирів. Так, у жировій тканині його питома вага складає 50% по відношенню до гліколізу, у печінці – 2,5 – 3 і в м'язовій тканині – 0,3%. Припускають, що останні стадії пентозного циклу забезпечують жирові клітини гліцерином, який утворюється з 3-фосфогліцеринового альдегіду.
Окрім описаних вище шляхів перетворення вуглеводів, в тканинах тваринних організмів були знайдені інші цикли, зокрема, перетворення моносахаридів шляхом їх з'єднання з піримідиновими основами і т.д.
Співвідношення між аеробним і анаеробним процесами перетворення вуглеводів в організмі. Основним шляхом перетворення вуглеводів у тканинах організму є поєднання анаеробного перетворення (гліколіз і глікогеноліз) і аеробного окислення за циклом трикарбонових кислот. Ці фази перетворення вуглеводів мають багато спільного. Обидва процеси розпочинаються з утворення фосфорних ефірів глюкози. Крім того, в обох випадках утворюються однакові проміжні продукти (3-фосфогліцериновий альдегід, фосфогліцеринова і фосфоенолпіровиноградна кислота, тощо). Центральною спільною ланкою, яка об'єднує ці процеси, є піровиноградна кислота, наступне перетворення якої залежно від умов може проходити як аеробним, так і анаеробним шляхом. Спряження процесів гліколізу і тканинного дихання зумовлено також тим, що багато реакцій гліколізу та аеробного перетворення вуглеводів каталізуються одними і тими самими ферментами і коферментами – зокрема НАД+, НАДФ+, ФМН, ФАД та деякими іншими.
Важливим шляхом регуляції співвідношення і зв'язку між анаеробною й аеробною фазами перетворення вуглеводів є ефект, виявлений Л. Пастером і названий на честь дослідника ефектом Пастера. Суть його полягає в тому, що під впливом кисню (за аеробних умов) анаеробне перетворення вуглеводів пригнічується. Отже, при наявності кисню анаеробний процес (гліколіз або спиртове бродіння) замінюється енергетично більш економним для клітини процесом перетворення вуглеводів – аеробним. За цих умов при менших витратах субстрату (глюкози) організм отримує значно більшу кількість енергії. Тому при наявності кисню значно зменшується використання субстрату – глюкози.
Дослідженнями O.В. Палладіна, В.O. Енгельгардта та інших учених встановлено, що основною причиною, яка зумовлює пастерівський ефект є, очевидно, своєрідна конкуренція між ферментними системами, які забезпечують аеробне й анаеробне перетворення вуглеводів, та захоплення неорганічного фосфату, необхідного для утворення АТФ.
У процесі обміну АДФ легко переходить із гіалоплазми в мітохондрії, де за аеробних умов інтенсивно використовується для синтезу АТФ, тобто ферментні системи аеробного перетворення вуглеводів ефективніше використовують неорганічний фосфат і АДФ. Зменшення вмісту АДФ у гіалоплазмі зумовлює гальмування гліколізу. Однак взаємозв'язок анаеробного й аеробного перетворення вуглеводів, очевидно, вивчено ще не повністю. Відомо, що в деяких органах і тканинах (сітківка ока, лейкоцити, ембріональна тканина, злоякісні пухлини) гліколіз досить ефективно проходить і за аеробних умов.
За певних умов гліколіз здатний пригнічувати аеробне перетворення вуглеводів. Пригнічення процесу дихання гліколізом дістало назву ефекту Кребтрі. Цей ефект найчастіше спостерігається при високих концентраціях глюкози, коли резерви АТФ досить швидко використовуються для синтезу гексозофосфорних ефірів – глюкозо-6-фосфату і фруктозо-1,6-дифосфату. При цьому вміст АТФ у гіалоплазмі значно зменшується, що призводить до переходу частини її з мітохондрій, де виникає дефіцит макроергів і пригнічується аеробний процес. Послабити ефект Кребтрі можна шляхом додаткового введення АТФ, що сприятиме активуванню процесів фосфорилювання глюкози. Глюкозо-6-фосфат стимулює і гліколіз, і дихання, тому залежно від умов у більшій або меншій мірі виявляються ефект Пастера або ефект Кребтрі.
Біосинтез дисахаридів
Важливими представниками дисахаридів є сахароза, лактоза, мальтоза та деякі інші. Біосинтез їх здійснюється в основному за реакціями трансглікозування. При цьому процес перенесення глікозильного залишку на один моносахарид проходить з фосфорного ефіру другого моносахариду. Реакція каталізується ферментом – специфічною глікозилтрансферазою. За приклад можна взяти синтез – сахарози:

Фермент, який каталізує цю реакцію, виділений з бактерій. Очевидно, в них так відбувається синтез сахарози.
Синтез сахарози може проходити й іншим способом, зокрема шляхом взаємодії уридиндифосфоглюкози з фруктозою. Реакцію каталізує фермент сахарозосинтаза:
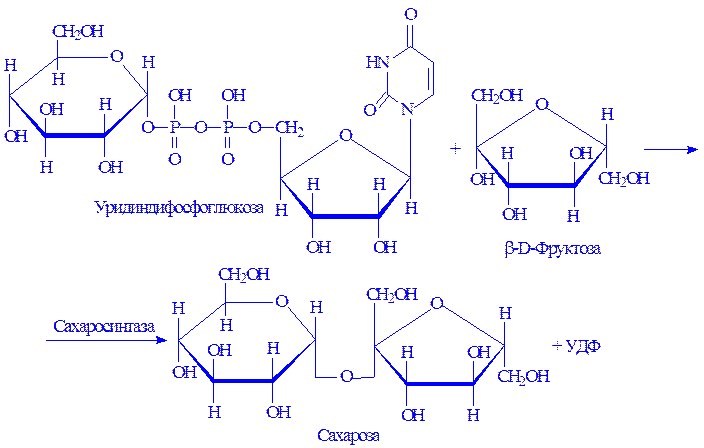
Так синтезується сахароза в рослинах.
Розглянемо біосинтез іншого дисахариду – лактози, яка складається з двох моноз – глюкози і галактози. Він проходить у кілька стадій. Спочатку галактозо-1-фосфат взаємодіє з уридиндифосфоглюкозою. При цьому утворюється уридиндифосфогалактоза і глюкозо-1-фосфат. Далі сполуки, що утворились, взаємодіють між собою, при цьому синтезуються лактоза та виділяється уридинтрифосфат. Першу стадію реакції каталізує фермент уридинтрансфераза, другу – глікозилтрансфераза:

Кінцевий обмін
Основними кінцевими продуктами обміну вуглеводів є вода і вуглекислий газ. Вода виділяється з сечею, потом, каловими масами, повітрям, що видихається. Вуглекислий газ виділяється легенями з повітрям, що видихається. В сечі міститься деяка кількість глюкуронової кислоти, яка, знешкоджуючи отруйні речовини, утворює парні сполуки. В калі містяться неперетравлені через якісь причини або важко переварювані (лігнін, клітковина) вуглеводи.

