




Общие соображения.
В каждом глазу около 125 миллионов палочек и колбочек, но всего 1 миллион ганглиозных клеток. Ввиду такого различия возникает вопрос: как при такой конвергенции может сохраняться детальная зрительная информация? Изучение связей между клетками сетчатки может помочь разрешить эту проблему. Существуют два пути информационного потока через сетчатку:
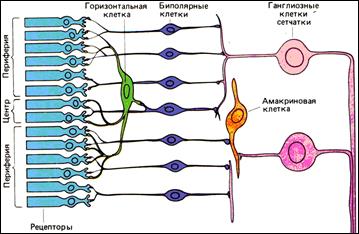 - Прямой путь, идущий от фоторецепторов к биполярным и далее к ганглиозным клеткам. Прямой путь весьма специфичен, или компактен, в том смысле, что одна биполярная клетка имеет входы лишь от одного рецептора или от сравнительно небольшого их числа, а одна ганглиозная клетка – от одного или сравнительно немногих биполяров.
- Прямой путь, идущий от фоторецепторов к биполярным и далее к ганглиозным клеткам. Прямой путь весьма специфичен, или компактен, в том смысле, что одна биполярная клетка имеет входы лишь от одного рецептора или от сравнительно небольшого их числа, а одна ганглиозная клетка – от одного или сравнительно немногих биполяров.
- Непрямой путь, при котором между рецепторами и биполярами могут быть включены еще горизонтальные клетки, а между биполярами и ганглиозными клетками – амакриновые клетки, более «размыт», или диффузен, благодаря более широким боковым связям.
Общая площадь, занятая рецепторами, связанными с одной ганглиозной клеткой по прямому и непрямому путям (до 1мм2) является рецептивным полем ганглиозной клетки – областью сетчатки, световая стимуляция которой может влиять на электрическую активность данной ганглиозной клетки.
Эта общая схема верна для всей сетчатки, но в деталях связей имеются большие различия между центральной ямкой, куда проецируется направление взгляда и где наша способность видеть тонкие детали максимальна, и периферией сетчатки, где острота зрения резко снижается. В центральной ямке или около нее на прямом пути, как правило, одна колбочка связана с одной биполярной клеткой, а один биполяр – с одной ганглиозной клеткой. При переходе от центральной ямки к периферии сеть прямых путей от рецепторов к ганглиозным клеткам становится совершенно иной: все больше рецепторов конвергируют на биполярах, а биполяров – на ганглиозных клетках. Эта высокая степень конвергенции, которую мы видим в большей части сетчатки, вместе с весьма компактными путями в самом центре и около него позволяют понять, почему, несмотря на отношение 125:1 между числом рецепторов и числом волокон зрительного нерва, некоторая часть сетчатки (ее центр) может все-таки обеспечивать острое зрение.
Слой биполяров.
Биполярная клетка имеет единственный дендрит, по которому получает сигналы от рецептора. Он либо образует синапс с одним рецептором (всегда с колбочкой) – меньшинство, либо расщепляется на веточки, образующие синапсы более чем с одним рецептором – большинство. А если учесть, что многие биполяры получают сигналы и от рецепторов через горизонтальные клетки, то становится очевидным наличие рецептивных полей и у биполяров.
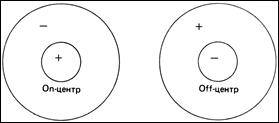 Если с одним биполяром связаны два или несколько рецепторов, они совместно занимают сравнительно малый участок сетчатки и составляют центр рецептивного поля. Периферию же рецептивного поля определяет непрямой путь от более обширной области рецепторов, связанных с биполярами через горизонтальные клетки. Таким образом биполярные клетки имеют рецептивные поля с центром и периферией. Самым удивительным является то, что воздействие света на центр и периферию рецептивного поля биполяра прямо противоположно, а сами биполяры разделяются на два типа: on- биполяры (с on- центром и off- периферией) и off- биполяры (с off- центром и on- периферией). «Оn» – включено, «off» – выключено.
Если с одним биполяром связаны два или несколько рецепторов, они совместно занимают сравнительно малый участок сетчатки и составляют центр рецептивного поля. Периферию же рецептивного поля определяет непрямой путь от более обширной области рецепторов, связанных с биполярами через горизонтальные клетки. Таким образом биполярные клетки имеют рецептивные поля с центром и периферией. Самым удивительным является то, что воздействие света на центр и периферию рецептивного поля биполяра прямо противоположно, а сами биполяры разделяются на два типа: on- биполяры (с on- центром и off- периферией) и off- биполяры (с off- центром и on- периферией). «Оn» – включено, «off» – выключено.
Оn- реакция подразумевает деполяризацию мембраны биполяра в ответ на световой стимул фоторецепторов и соответственно усиленное выделение медиатора в выходных синапсах, off-реакция противоположна – гиперполяризация и уменьшение выброса медиатора в ответ на освещение фоторецепторов.
Очевидно, что подобные реакции зависят от того, каковы синапсы между рецепторами и биполярными клетками, рецепторами и горизонтальными клетками, горизонтальными клетками и биполярами – возбуждающие они, тормозные или обоих типов.
Синапсы между рецепторами и биполярами (прямой путь - центр рецептивного поля).
Рецепторы активны в темноте, а свет, вызывая гиперполяризацию, уменьшает их активность, следовательно:
- если синапс возбуждающий, биполяр будет активироваться в темноте, а инактивироваться на свету – off-реакция;
- если же синапс тормозной, биполяр в темноте тормозится, а свет, выключая рецептор, снимает это торможение и биполярная клетка (в этом случае – спонтанно активный нейрон) активируется – оn- реакция.
Рецептор высвобождает лишь один тип медиатора, следовательно, является ли рецепторно-биполярный синапс возбуждающим или тормозным, зависит от типа каналов в постсинаптической мембране биполярной клетки.
Синапсы горизонтальных клеток.
О механизмах работы горизонтальных клеток известно значительно меньше, однако очевидно следующее: там, где горизонтальные клетки непосредственно контактируют с биполярами, синапсы с on-биполярами должны быть возбуждающими (ввиду тормозящего влияния света на периферию), а с off-биполярами – тормозными. Если же влияние осуществляется через рецепторы, то все синапсы должны быть тормозными.
Наличие в рецептивных полях центра и периферии с противоположной реакцией на освещение является структурно-физиологической основой описанного ранее латерального торможения.
Этот принцип конвергенции сигналов и латерального торможения действует и на следующем уровне обработки информации в сетчатке (ганглиозные и амакриновые клетки)
РП горизонтальных клеток, как правило, сравнительно велики. Функционально они связаны друг с другом. Некоторые горизонтальные клетки (L–muпа) гиперполяризуются при попадании света на их РП независимо от его спектрального состава; другие при освещении либо деполяризуются, либо гиперполяризуются в зависимости от того, какой части спектра соответствует стимул (цветоспецифичные красно–зеленые или сине-желтые горизонтальные клетки).
Слой ганглиозных клеток
Структура рецептивных полей (РП) ганглиозных клеток является аналогичной – существуют ганглиозные клетки с on-центром и off-периферией и с off-центром и on-периферией, однако из-за усиливающейся конвергенции размеры РП увеличиваются.
Как правило, несколько on- и off-биполяров конвергируют на одну ганглиозную клетку (прямой путь). Значительно больше биполярных клеток связаны с ганглиозной через амакриновые клетки, формируяпериферию ее с рецептивного поля (непрямой путь).
При этом ганглиозные клетки с off-центром на прямом пути (центр РП) возбуждаются off-биполярами и тормозятся оn-биполярами. На непрямом пути (периферия РП) – латерально тормозятся off-амакриновыми клетками и возбуждаются амакриновыми клетками, возбуждаемыми оn-биполярами.
Реакция ганглиозных клеток на свет.
 Ганглиозные клетки с оn-центром деполяризуются в ответ на освещение центра рецептивного поля, и эта деполяризация в аксонном холмике преобразуется в последовательность потенциалов действия. С другой стороны, как освещение периферии РП, так и «выключение света» в центре РП вызываютгиперполяризацию их мембранного потенциала и, следовательно, кратковременное торможение нейронной активности.
Ганглиозные клетки с оn-центром деполяризуются в ответ на освещение центра рецептивного поля, и эта деполяризация в аксонном холмике преобразуется в последовательность потенциалов действия. С другой стороны, как освещение периферии РП, так и «выключение света» в центре РП вызываютгиперполяризацию их мембранного потенциала и, следовательно, кратковременное торможение нейронной активности.
Если стимулировать одиночную ганглиозную клетку с on-центром все более крупными световыми пятнами, реакция будет постепенно усиливаться вплоть до пятна с величиной около 1 градуса. Это совпадает с величиной центра. Дальнейшее увеличение пятна ведет к уменьшению реакции, так как при этом пятно начинает захватывать антагонистическую периферию. При размерах пятна более 3 градусов реакция перестает уменьшаться, поэтому 3 градуса — это поперечник всего рецептивного поля, включая центр и периферию.
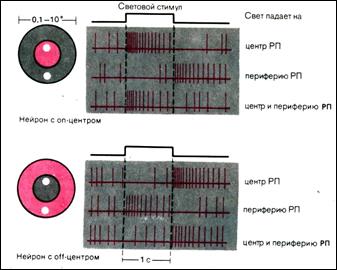 Рецептивные поляганглиозных клеток с off–центром функционально противоположны только что описанным. Освещение их центра приводит к «прямому» торможению, а ослабление падающего света– к «off–активации». Освещение периферии РП ведет к латеральной активации, а «выключение света» на периферии – к кратковременному «латеральному» торможению частоты импульсации нейрона.
Рецептивные поляганглиозных клеток с off–центром функционально противоположны только что описанным. Освещение их центра приводит к «прямому» торможению, а ослабление падающего света– к «off–активации». Освещение периферии РП ведет к латеральной активации, а «выключение света» на периферии – к кратковременному «латеральному» торможению частоты импульсации нейрона.
Если центр и периферия РП освещаютсяодновременно, доминирует реакция центра. Однако при этом реакция слабее, чем при освещении только центра РП, поскольку возбуждение или торможение центра и периферии РП суммируются.
Функциональная организация рецептивных полей
ганглиозных клеток сетчатки млекопитающих.
При анализе рецептивных полей небольшие пятна света (показаны белым) проецировались либо на центр, либо на периферию РП. Световые стимулы вызывают разный ответ у нейронов с оn– и off–центром. Когда обе части РП освещены одновременно, возбуждающий и тормозный процессы, связанные с освещением центра и периферии, суммируются. Однако преобладает ответ, вызываемый стимуляцией центра РП
Ганглиозные клетки on–off–типа обычно дают короткий оn–ответ на стационарный световой стимул и короткую off–реакцию на ослабление света. В этот класс клеток входят, например,нейроны, чувствительные к движению, особенно хорошо воспринимающие перемещение через их рецептивное поле границы света и темноты. При этом степень возбуждения зависит от угловой скорости стимула.
Еще одна классификация ганглиозных клеток основана наскорости проведения сигналов по их аксонам. У большинства крупных ганглиозных клеток со сравнительно толстыми миелинизированными аксонами она высока. На освещение рецептивного поля они дают короткий «фазический» ответ (нейроны I класса латентности, илиY–нейроны ). Гораздо многочисленнее мелкие ганглиозные клетки с более тонкими миелинизированными аксонами, которые на освещение центра РП реагируют «тоническим» возбуждением или торможением (нейроны II класса латентности, илиХ–нейроны ). Среди Х– и Y–нейронов клетки с on– и off–центром одинаково часты. Наконец, в сетчатке есть ганглиозные клетки с тонкими, лишь слегка миелинизированными аксонами (III класса латентности, или W–нейроны). К этому классу в основном и относятся on–off–нейроны, чувствительные к движению.
Даже если не рассматривать цветоспецифичные реакции ганглиозных клеток сетчатки, очевидно, что она представляет собой сложную нейронную систему. Уже внутри нее самой изображение, активирующее входной слой рецепторов, преобразуется в несколько типов возбуждения различных ганглиозных клеток.
Итак:
1. Рецептивные поля большинства ганглиозных клеток, как и биполяров организованыантагонистически, т.е. световой стимул вызывает ихдиаметрально противоположную реакцию в зависимости от того, попадает он на периферию или в центр рецептивного поля.
2. Сетчатка содержит примерно равные количества ганглиозныхклеток c on- и off -центрами.
3. Такая пространственная организация РП обеспечивается двумя способами: конвергенцией сигналов от нескольких рецепторов на одной клетке и опосредованными тормозными сигналами с периферии рецептивного поля, доходящими до биполяров через горизонтальные клетки, а до ганглиозных – через амакриновые клетки (латеральное торможение).
4. Возбуждение фоторецепторов фотонами передается в ЦНС по двум независимым нейронным «каналам»: через оn-систему, активируемую, когда световая стимуляция в центре РП сильнее, чем на периферии, и через off-систему, активируемую при уменьшении освещения.
5. Зрительный нерв образован аксонами ганглиозных клеток с on- и off -центрами.
Задание
1. В ходе изучения работы сетчатки исследователи были поражены тем, что магниевая вспышка, направленная прямо в глаз животного, вызывала столь слабые реакции, регистрируемые на зрительном нерве или не вызывала их вовсе. Казалось бы, следовало ожидать, что освещение всех рецепторов, гарантируемое при такой вспышке, будет наиболее сильным, а не слабым стимулом. Объясните этот парадокс.
2. Изобразите четыре разных варианта нейронных сетей, ведущих от фоторецепторов к ганглиозным клеткам.
Значение рецептивных полей с центром и периферией
Почему в ходе эволюции сформировались столь сложные рецептивные поля с центральными и периферическими зонами? В чем адаптивность подобной организации сетчатки? Иными словами, какую пользу это приносит животному?
Сообщения, которые глаз посылает мозгу, могут быть очень мало связаны с абсолютной освещенностью сетчатки, так как ганглиозные клетки сетчатки слабо реагируют на изменения силы рассеянного света. О чем действительно сигнализирует клетка, так это о результате сравнения количества света, падающего на определенный участок сетчатки, с его средним количеством в ближайшем окружении.
Это можно проиллюстрировать следующим экспериментом:
1) Картируем рецептивное поле одной ганглиозной клетки. Пусть это будет клетка с on-центром.
2) Равномерно осветив экран слабым постоянным фоновым светом, начнем включать и выключать пятнышко, точно заполняющее центр поля, начиная со столь слабого света, что его еще нельзя увидеть, и постепенно повышая интенсивность.
3) При некоторой яркости пятна мы начнем обнаруживать реакцию клетки, именно в тот момент, когда пятно станет различимо и невооруженным глазом. Измерив интенсивность фона и пятнышка фотометром, мы выясним, что пятнышко приблизительно на 2% ярче фона.
4) Повторим всю процедуру, начиная с фонового света в пять раз более яркого, чем в первом эксперименте.
5) В момент, когда мы снова начнем обнаруживать реакции клетки и сами различать пятно, измерим стимулирующий свет. Он тоже в пять раз ярче предыдущего, т. е. пятнышко снова на 2% ярче фона.
Вывод таков, что как для нас, так и для отдельной клетки, существенна относительная освещенность пятнышка и фона. Мы способны производить такое сравнение при различии всего в 2% – в точности так, как это могут делать наиболее чувствительные ганглиозные клетки в сетчатке обезьяны.
Ганглиозная клетка, если учесть ее физиологию, может передавать информацию только о границах пятна; внутренность его мы видим как однородную, поскольку ганглиозные клетки с полями, находящимися внутри пятна, не сообщают о локальных различиях в освещенности. Видеть большое пятно с помощью только тех клеток, поля которых лежат на его границах (не нуждаясь в участии остальных клеток с центрами, распределенными по всему пятну), — это более экономный способ..
Наряду с эффективностью эта система обладает еще одним важным преимуществом. Большинство объектов мы видим благодаря отраженному свету таких источников, как солнце или электрическая лампочка. Несмотря на изменения освещенности, создаваемой этими источниками, наша зрительная система сохраняет удивительную способность к неизменному восприятию объектов. Ганглиозная клетка сетчатки работает именно так, что это становится возможным.
Рассмотрим следующий пример: печатный текст выглядит примерно одинаково – белая бумага, черные буквы, – читаем ли мы ее в полумраке комнаты или на пляже в солнечный день. Предположим, что в обоих случаях мы измерим входящий в наши глаза свет отраженный от белой бумаги и от одной из черных букв заголовка. Снаружи свет в 20 раз ярче, чем в комнате, а черные буквы отражают приблизительно десятую долю света, отражаемого белой бумагой. Получаем следующие данные:
| На открытом воздухе | В комнате | |
| Белая бумага | 6,0 | |
| Черные буквы | 0,6 |
Они показывают, что черная буква на открытом воздухе посылает в наши глаза вдвое больше света, чем белая бумага при комнатном освещении! Отсюда ясно, что восприятие черного и белого не определяется самим по себе количеством отражаемого объектом света. Существенно то, каково это количество в сравнении с окружающим фоном.
Экран выключенного телевизора в нормально освещенной комнате выглядит серовато-белым. Мы, однако, хорошо знаем, что телевизор может дать нам ощущение насыщенного черного цвета. Самый темный участок изображения посылает в наши глаза по меньшей мере столько же света, что и при выключенном телевизоре. Отсюда следует, что «черное» и «белое» — не просто физические понятия: это биологические термины, они отражают результат вычислений, производимых нашей сетчаткой и мозгом при обработке воспринимаемой картины.
Все высказанные соображения о черном и белом, применимы и к цвету. Видимый цвет объекта определяется не только приходящим от него светом, но также и светом, приходящим от всего остального окружения. В результате то, что мы видим, становится независимым не только от интенсивности света, но и от его спектрального состава. И опять-таки это способствует постоянству восприятия видимой картины несмотря на заметные различия в яркости и спектральном составе падающего на нее света.

