




Основным источником энергии в организме является глюкоза. Концентрация глюкозы в крови строго контролируется. Глюкоза необходима, прежде всего, как источник энергии для мозга. Одна из основных функций печени состоит в поддержании концентрации глюкозы в крови на уровне около 4,5 ммоль/л (округленные значения для лучшего запоминания 3,33-5,55 ммоль/л). Если уровень глюкозы в крови снижается, развиваются нарушения метаболизма в центральной нервной системе, которые могут приводить к коме, судорогам и при выраженной гипогликемии – к смерти. В поддержании уровня глюкозы в крови принимают участие инсулин, глюкагон, адреналин и глюкокортикоиды, которые регулируют метаболизм во многих тканях, но особенно в печени, мышцах и жировой ткани.
Инсулин
1. Синтез инсулина. Синтез инсулина происходит в b-клетках островков Лангерганса поджелудочной железы.
Ген человеческого инсулина локализован в коротком плече 11 хромосомы. Инсулин синтезируется на рибосомах шероховатого эндоплазматического ретикулума в виде препроинсулина (М.м. 11500), который на N-конце содержит сигнальный пептид, состоящий из 16 аминокислот и направляющий пептидную цепь в просвет эндоплазматического ретикулума. В ЭПР сигнальный пептид отделяется и после замыкания дисульфидных связей образуется проинсулин (М.м. 9000). Биологическая активность проинсулина составляет 5% биологической активности инсулина. Проинсулин поступает в аппарат Гольджи, где в секреторных везикулах происходит отщепление эквимолярного количества С-пептида и образуется зрелый инсулин, который сохраняется в форме цинксодержащего гексамера вплоть до секреции. Мембраны секреторных пузырьков (гранул) в процессе секреции сливаются с плазматической мембраной клетки, а их содержимое высвобождается во внеклеточное пространство. Определение в крови концентрации С-пептида может быть использовано для определения функции поджелудочной железы (при введении экзогенного инсулина или при невозможности прямого определения инсулина в сыворотке крови из-за наличия противоинсулиновых антител).
2. Структура инсулина. Молекула инсулина – полипептид, состоящий из 2-х цепей, А цепь (21 аминокислотный остаток) и В цепь (30 аминокислотных остатков). Цепи связанны между собой дисульфидными мостиками. Дисульфидные мостики локализованы между аминокислотными остатками А7-В7 и А20-В19. Третий дисульфидный мостик связывает между собой 6 и 11 аминокислотные остатки А цепи. Локализация всех трех дисульфидных мостиков постоянна.
Выделяют 3 консервативных участка в молекуле инсулина; 1) положение 3-х дисульфидных связей; 2) гидрофобные остатки в С-концевом участке В-цепи и 3) С- и N-концевые участки А-цепи. Наиболее схожи по своему строению инсулины человека, свиньи (отличие по 1 аминокислоте) и быка (3 аминокислоты), что позволяет их использовать в качестве препаратов заместительной терапии при сахарном диабете.
Поджелудочная железа человека секретирует до 40-50 ед. инсулина в сутки, что соответствует 15-20% общего количества гормона в железе.
3. Регуляция синтеза инсулина. Повышение концентрации глюкозы в крови – главный физиологический стимул секреции инсулина. Пороговой для секреции инсулина является концентрация глюкозы натощак 80-100 мг/дл (>5,5 ммоль/л), а максимальная секреция наблюдается при концентрации глюкозы 300-500 мг/дл (15-20 ммоль/л). Кроме того, стимулируют синтез и секрецию инсулина аминокислоты лейцин, глюкагон, гормон роста, кортизол, плацентарный лактоген, эстрогены и прогестерон. Ингибируется синтез инсулина адреналином.
4. Деградация инсулина. В крови инсулин не имеет белков-переносчиков. Период полужизни 3-5 минут. Катаболизм инсулина происходят в основном в печени, почках и плаценте. Около 50% инсулина метаболизируется за один пассаж крови через печень. В деградации инсулина участвуют 2 ферментативные системы: 1) инсулин-специфическая протеиназа, расщепляющая инсулин до аминокислот и 2) глутатион-инсулин-трансгидрогеназа, который восстанавливает дисульфидные мостики.
5. Формы инсулина в крови. Различают 3 формы инсулина в крови: 1) свободная форма инсулина – способствует утилизации глюкозы жировой и мышечной тканью; 2) связанная с белками форма инсулина – влияет только на жировую ткань; 3) форма А – промежуточная форма инсулина, появляется в крови в ответ на быструю, срочную потребность организма в инсулине.
5. Механизм действия инсулина. По механизму действия инсулин относится к гормонам со смешанным механизмом действия. Вначале инсулин связывается со специфическим гликопротеиновым рецептором, который содержит много гликозильных остатков на поверхности клетки-мишени. Удаление сиаловых кислот и галактозы снижает способность рецептора связывать инсулин и активность гормона.
Рецептор инсулина состоит из 2-х a и 2-х b-субъединиц, соединенных дисульфидными мостиками. a-Субъединица расположена вне клетки и осуществляет связывание инсулина. b-субъединица обладает тирозинкиназной активностью и содержит участок аутофосфорилирования.
В результате связывания инсулина с рецептором происходит изменение конформации рецептора, гормон-рецепторный комплекс проникает в цитозоль путем эндоцитоза (интернализация), распадается и генерируется сигнал внутри клетки. Рецепторы могут подвергаться протеолизу или повторному процессингу и вновь встраиваться в мембрану. В качестве внутриклеточных посредников выступает сам инсулин, ионы кальция, циклические нуклеотиды, продукты распада фосфатидилинозитола, пептиды мембран.
Различные эффекты инсулина делятся на 1) быстрые, которые проявляются через несколько секунд или минут (деполяризация мембран, транспорт глюкозы и ионов, фосфорилирование белков, активация или ингибирование ферментов, синтез РНК) и 2) медленные – от нескольких часов до суток (синтез белка, ДНК, пролиферация клеток).
6. Метаболические эффекты инсулина.
Все органы делятся на инсулинчувствительные (мышечная, жировая ткани и частично печень) и инсулиннечувствительные (нервная ткань, эритроциты).
Основное биологическое значение инсулина – превращение избытка глюкозы крови в две резервные формы – гликоген (печень и мышцы) и триацилглицеролы (жировая ткань).
Обмен углеводов. Инсулин – единственный гормон, который снижает уровень глюкозы в крови по следующим механизмам.
1. Инсулин повышает проницаемость мембран мышечной и жировой тканей для глюкозы, увеличивая количество переносчиков для глюкозы и транслокацию их из цитозоля в мембрану. Гепатоциты хорошо проницаемы для глюкозы и инсулин способствует задержке глюкозы в клетках печени, стимулируя активность глюкокиназы и ингибируя глюкозо-6-фосфатазу. В результате быстро протекающего фосфорилирования концентрация свободной глюкозы в гепатоцитах поддерживается на очень низком уровне, что способствует ее проникновению в клетки по градиенту концентрации.
2. Инсулин влияет на внутриклеточную утилизацию глюкозы следующими путями: 1) ~50% поглощенной глюкозы превращается в энергию (гликолиз); 2) 30-40% - в жиры и 3) ~ 10% - в гликоген.
3. Инсулин усиливает интенсивность гликолиза в печени, повышая активность ферментов глюкокиназы, фосфофруктокиназы и пируваткиназы. Более интенсивный гликолиз способствует более активной утилизации глюкозы и, следовательно, способствует снижению выхода глюкозы из клетки.
4. В печени и мышцах инсулин стимулирует синтез гликогена путем ингибирования аденилатциклазы и активации фосфодиэстеразы. В результате концентрация цАМФ уменьшается, что приводит к активации гликогенсинтазы и ингибированию фосфорилазы.
5. Инсулин ингибирует глюконеогенез в результате уменьшения концентрации фосфоенолпируваткарбоксикиназы (ингибирование транскрипции генов и синтез мРНК).
Обмен липидов.
1. Инсулин стимулирует липогенез в печени и жировой ткани путем:
а) увеличения концентрации ацетил КоА и НАДФН+Н+, необходимых для синтеза жирных кислот в результате активации пируватдегидрогеназного полиферментного комплекса и пентозофосфатного пути распада глюкозы;
б) активации фермента ацетил-КоА-карбоксилазы, катализирующего превращение ацетил-КоА в малонил-КоА;
в) активации полиферментного комплекса синтазы высших жирных кислот путем дефосфорилирования;
г) увеличения притока глицерола, необходимого для синтеза триацилглицеролов.
2. В печени и жировой ткани инсулин через снижение концентрации цАМФ ингибирует гормончувствительную липазу и липолиз.
3. Инсулин ингибирует синтез кетоновых тел в печени.
4. Инсулин стимулирует синтез ЛПОНП, которые транспортируют триацилглицеролы из печени в периферические ткани.
Обмен белков. Инсулин оказывает анаболический эффект на обмен белков, поскольку он стимулирует синтез и тормозит распад белков. Инсулин стимулирует поступление нейтральных аминокислот в мышечную ткань. Влияние инсулина на синтез белков в скелетных и сердечной мышцах проявляется, вероятно, на уровне трансляции мРНК.
Пролиферация клеток. Инсулин стимулирует пролиферацию клеток в клеточных культурах и возможно участвует в регуляции роста in vivo.
Нарушение метаболизма инсулина. При недостатке инсулина развивается сахарный диабет. Примерно у 90% больных диабетом наблюдается инсулиннезависимый сахарный диабет II типа. Характерен для людей зрелого возраста. Для таких больных типичны ожирение, повышенное содержание в плазме инсулина и снижение количества инсулиновых рецепторов. У 10% больных наблюдается диабет I типа (инсулинзависимый, ювенильный), который остро начинается в юношеском возрасте. Обусловлен поражением поджелудочной железы различными факторами и снижением количества инсулина в крови. Разрушение β-клеток может быть вызвана ксенобиотиками и лекарственными препаратами, вирусами, аутоиммунными процессами.
Метаболические изменения при диабете. Основными признаками недостаточности инсулина являются: гипергликемия, кетоацидоз и гипертриацилглицеролемия. Гипергликемия обусловлена снижением утилизации глюкозы периферическими тканями и повышенной продукцией глюкозы за счет активации глюконеогенеза и гликогенолиза. Когда концентрация глюкозы превышает порог реабсорбции, глюкоза экскретируется с мочой (глюкозурия). Повышенная мобилизация жирных кислот приводит к повышенной продукции кетоновых тел и развитию кетоацидоза. При диабете повышаются превращение жирных кислот в триацилглицеролы и секреция ЛПОНП и хиломикронов, что приводит к увеличению их концентрации в крови.
На протяжении многих лет ведется поиск гипогликемических препаратов с новыми механизмами действия: секретогенов, сенситайзеров инсулина, ингибиторов скорости продукции глюкозы печенью и др. Одним из перспективных направлений является поиск модуляторов инкретинов: GIP и GLP, которые регулируют активность секреции β-клеток путем оптимизации синтеза инсулина и способствуют пролиферации β-клеток, неогенезу островков Лангерганса, а также препятствуют апоптозу β-клеток.
Снижение чувствительности рецепторов к инсулину лежит в основе инсулинорезистентности. Инсулинорезистентность является ведущей причиной развития метаболического синдрома у каждого 4-6 жителя Земли. Метаболический синдром – это прогрессирующее состояние предболезни у людей постиндустриального общества. В самом общем виде метаболический синдром включает пять важнейших признаков: избыточная масса тела или ожирение, артериальная гипертензия, гипергликемия натощак, снижение концентрац ХС ЛПВП и повышение концентрации триацилглицеролов в плазме крови.
Глюкагон
Глюкагон – полипептид, состоящий из 29 аминокислот (м.м. 3500 Да). Был выделен в 1953 г. Штраубом. Основным местом синтеза глюкагона являются a-клетки островкового аппарата поджелудочной железы. Период полужизни около 15-20 минут. Инактивация гормона происходит в печени путем протеолиза.
Регуляция секреции. Через несколько часов после приема пищи, даже при отсутствии значительной физической активности или стресса, уровень глюкозы в крови начинает падать вследствие окисления глюкозы в мозге и других тканях. Снижение уровня глюкозы в крови вызывает секрецию глюкагона и ингибирует секрецию инсулина.
Метаболические эффекты глюкагона. Глюкагон – непроникающий гормон, рецепторы находятся на поверхности клеток. Основные органы-мишени – печень и жировая ткань, в меньшей степени – мышцы. Механизм действия гормона реализуется по аденилатциклазному каскадному пути.
Обмен углеводов.
1. Глюкагон является основным гормоном, который повышает уровень глюкозы в крови.
2. В печени глюкагон активирует расщепление гликогена (гликогенолиз) путем активации гликогенфосфорилазы и инактивации гликогенсинтазы. Регуляция активности обоих ферментов осуществляется путем фосфорилирования через цАМФ. Глюкагон не влияет на распад гликогена в мышцах.
3. Глюкагон ингибирует гликолиз в печени и активирует глюконеогенез. Эти эффекты обусловлены снижением уровня фруктозо-2,6-бисфосфата, который является аллостерическим ингибитором фруктозо-1,6-бисфосфатазы и активатором фосфофруктокиназы. Глюкагон также ингибирует гликолитический фермент пируваткиназу, блокируя превращение фосфоенолпирувата в пируват и окисление пирувата в ЦТК.
Стимуляция глюкагоном распада гликогена в печени, ингибирование гликолиза и активация глюконеогенеза способствует поступлению глюкозы из печени в кровь и восстановление ее уровня до нормальных значений.
Обмен липидов.
1. В жировой ткани глюкагон активирует триацилглицероллипазу через цАМФ по аденилатциклазному механизму и увеличивает липолиз.
2. Образующиеся в большом количестве жирные кислоты транспортируются в печень и другие ткани, где используются в качестве источника энергии, сберегая глюкозу для мозга.
3. В печени жирные кислоты могут превращаться в кетоновые тела через ацетилКоА.
Обмен белков. Глюкагон активирует захват аминокислот печенью и стимулирует распад белков. Образованные аминокислоты используются для глюконеогенеза и после дезаминирования – для биосинтеза мочевины.
Гормоны мозгового вещества надпочечников (катехоламины)
В мозговом веществе надпочечников синтезируются норадреналин и адреналин. Роль этих гормонов заключается в адаптации к острым и хроническим стрессам.
Синтез катехоламинов происходит в хромаффинных клетках мозгового вещества надпочечников из аминокислоты тирозин. Свое название эти клетки получили потому, что содержат гранулы, окрашивающиеся под действием бихромата калия в красно-коричневый цвет. Главный продукт мозгового слоя надпочечников – адреналин, который составляет ~ 80% всех катехоламинов (1-3 мг/г адреналина и 0,2-0,6 мг/г норадреналина). Вне мозгового вещества адреналин не образуется. В отличие от него норадреналин, обнаруживаемый в органах, иннервируемых симпатическими нервами, образуется преимущественно in situ; остальная часть норадреналина образуется главным образом в окончаниях нервов и достигает своих мишеней с кровью.
Превращение тирозина в адреналин включает четыре последовательных этапа: 1) гидроксилирование кольца, 2) декарбоксилирование, 3) гидроксилирование боковой цепи, 4) N-метилирование. Благодаря метилированию адреналин приобрел способность циркулировать в кровеносном русле.
Запасание и секреция катехоламинов. В мозговом слое надпочечников содержатся хромаффинные гранулы – органеллы, способные к биосинтезу, поглощению, запасанию и секреции катехоламинов. Нервная стимуляция мозгового слоя надпочечников приводит к слиянию хромаффинных гранул с плазматической мембраной и, таким образом, обусловливает выброс норадреналина и адреналина путем экзоцитоза. Секретируемый надпочечниками адреналин попадает в печень и скелетные мышцы, но затем быстро метаболизируется. Катехоламины циркулируют в плазме в слабоассоциированном с альбумином виде. Период полужизни составляет 10-30 с. Концентрация адреналина в плазме крови составляет 2 нмоль/л. При стрессе количество катехоламинов увеличивается в 4-8 раз.
Метаболизм катехоламинов. Лишь очень небольшая часть адреналина (менее 5%) выделяется с мочой. Катехоламины быстро метаболизируются под действием катехол-О-метилтрансферазы (цитозольный фермент, обнаруживается во многих тканях) и моноаминооксидазы с образованием О-метилированных и дезаминированных продуктов, которые выводятся с мочой.
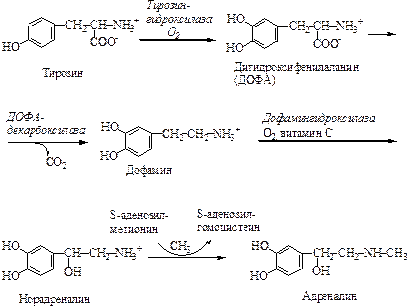
Механизм действия катехоламинов. Катехоламины действуют через 2 типа рецепторов: a-адренергические и b-адренергические. Каждый из них подразделяется на α1 и α2, β1 и β2. Адреналин связывается (и активирует) как с α-, так и с β-рецепторами, и поэтому его действие на ткань, содержащую рецепторы обоих классов, зависит от относительного сродства этих рецепторов к гормону. Норадреналин в физиологических концентрациях связывается главным образом с α-адренорецепторами.
При связывании адреналина с b-адренорецепторами активируется аденилатциклаза и увеличивается концентрация цАМФ. a-Адренорецепторы участвуют в процессах, ведущих к изменению внутриклеточной концентрации кальция или к изменению метаболизма фосфатидилинозитола.
Влияние на метаболизм: адреналин действует на мышцы, жировую ткань и печень.
Обмен углеводов. Адреналин активирует распад гликогена (гликогенолиз) в мышцах и в меньшей степени в печени, стимулирует синтез глюкозы в печени (глюконеогенез) и понижает поглощение глюкозы в мышцах и других тканях.
Адреналин вызывает повышение концентрации глюкозы в крови при остром стрессе для работы мозга и экстренного сокращения мышц.
Адреналин уменьшает секрецию инсулина и увеличивает секрецию глюкагона, что приводит к повышению концентрации глюкозы в крови, которая используется в качестве источника энергии для мозга.
Обмен липидов. В жировой ткани адреналин и норадреналин стимулируют распад триацилглицеролов за счет повышения содержания цАМФ и активации гормон-чувствительной липазы. Жирные кислоты используются в качестве источника энергии для мышечной активности и, кроме того, могут активировать глюконеогенез в печени.
Влияние на физиологические функции. Действуя на сердце, адреналин увеличивает минутный объем в результате повышения силы (инотропный эффект) и частоты (хронотропный эффект) сердечных сокращений и повышает артериальное давление, что обеспечивает повышенное поступление кислорода для окислительных процессов в тканях. Катехоламины вызывают расслабление гладкой мускулатуры бронхов, желудочно-кишечного тракта и кровеносных сосудов скелетных мышц. С другой стороны, катехоламины стимулируют сокращение гладкой мускулатуры сосудов кожи и почек.
Глюкокортикоиды
К стероидным гормонам относятся гормоны коры надпочечников (глюкокортикоиды и минералокортикоиды) и половые гормоны (эстрогены, прогестерон и тестостерон).
Синтез стероидных гормонов происходит по общей схеме и регулируется гормонами гипофиза. Стероидные гормоны образуются из холестерола, который поступает в периферическую эндокринную железу в составе ЛПНП или синтезируется внутриклеточно из ацетил-КоА. Значительная часть холестерола подвергается эстерификации и накапливается в цитоплазме в виде эфиров. При синтезе гормонов происходит активация холестеролэстеразы и образующийся свободный холестерол транспортируется в митохондрии, где превращается в общий промежуточный продукт – прегненолон. Прегненолон транспортируется в ЭПР, где через ряд промежуточных метаболитов превращается в стероидные гормоны.
Последовательность синтеза гормонов коры надпочечников представлена на рис. 10.3.
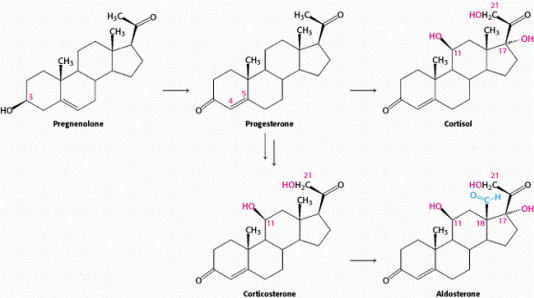
Рис. 10.3. Схема образования пргестерона, кортизола и альдостерона
Подписи: Прегненолон; Прогестерон; Кортизол; Кортикостерон; Альдостерон
Кортикотропин в коре надпочечников и лютропин в половых железах регулируют синтез стероидных гормонов: 1) увеличивают число рецепторов для ЛПНП на поверхности мембраны клеток; 2) активируют холестеролэстеразу и способствуют образованию свободного холестерола; 3) активируют ферменты превращения холестерола в прегненолон в митохондриях; 4) стимулирует распад углеводов и липидов, обеспечивая стероидогенез энергетическим и пластическим материалом.
В крови 90-95% стероидных гормонов связаны с белками.
Активны только свободные формы стероидных гормонов.
По обратной связи стероидные гормоны тормозят продукцию тропных гормонов гипофиза. Период полужизни стероидных гормонов 0,5-1,5 часа. В печени, почках и других тканях гормоны модифицируются (восстановление двойных связей, гидроксилирование и др.), что приводит к потере биологической активности. Повышение гидрофильности достигается образованием парных соединений (глюкуронаты, сульфаты, фосфаты), которые выводятся с мочой.
Гормоны коры надпочечников. Кора надпочечников взрослого человека состоит из трех четко различимых слоев, или зон. Субкапсулярная зона называется клубочковой зоной; она связана с продукцией минералокортикоидов. Следующей идет пучковая зона; в ней, а также в сетчатой зоне вырабатываются глюкокортикоиды и андрогены. Из ткани коры надпочечников выделено и получено в кристаллическом виде около 50 стероидов. Большинство из них – промежуточные продукты; только немногие секретируются в значительном количестве и совсем малое число обладает значительной гормональной активностью. В целом наблюдается перекрывание их биологической активности; так, все природные глюкокортикоиды проявляют минералокортикоидный эффект и, наоборот, минералокортикоиды обладают глюкокортикоидной активностью.
Глюкокортикоиды – стероиды, состоящие из 21 углеродного атома. В основе строение – циклопентанпергидрофенантреновая структура, в 3-м и 20-м положении – кето(оксо)-группа, двойная связь в 4,5 положении. Основной глюкокортикоид человека – кортизол имеет дополнительно в 11 и 17 положениях гидроксигруппы; у грызунов – кортикостерон, имеет дополнительно в 11 положении гидроксигруппу. Кортизон дополнительно в 11 положении имеет оксогруппу, а в 17 положении – гидроксигруппу.
Глюкокортикоиды играют важную роль в адаптации к сильным и продолжительным стрессам.
Регуляция синтеза и секреции глюкокортикоидов осуществляется кортиколиберином гипоталамуса и кортикотропином гипофиза. В сутки синтезируется 20-25 мг кортизола. Существует циркадный ритм выработки гормона. Увеличение синтеза происходит сразу после засыпания. Во время сна уровень кортизола продолжает возрастать, достигая пика вскоре после просыпания, затем постепенно падает до минимальных величин к концу дня и в ранние вечерние часы.
Транспорт в крови. Кортизол в плазме крови находится в связанной с белками и свободной форме. Активной является свободная форма гормона. Основной связывающий белок плазмы крови - a-глобулин – транскортин. Транскортин синтезируется в печени и синтез его стимулируется эстрогенами. Меньшее количество кортизола транспортируется альбуминами. Период полужизни кортизола ~ 1,5-2 ч, у кортикостерона - < 1 часа (т.к. он слабее связывается с транспортными белками).
Метаболизм и экскреция. Метаболизм глюкокортикоидов в основном происходит в печени и включает восстановление двойных связей в первом кольце и 3-оксогруппы. Восстановленные метаболиты в печени подвергаются конъюгации с глюкуроновой или серной кислотами, благодаря чему они становятся более гидрофильными и выводятся из организма. Около 70% конъюгированных стероидов экскретируется с мочой, 20% - с калом, остальное – через кожу. Небольшое количество кортизола при отщеплении боковой цепи превращается в 17-кетостероиды.
Органы-мишени: печень, почки, лимфоидная ткань (селезенка, лимфоузлы, лимфоциты, тимус и др.), соединительная ткань (кости, подкожная клетчатка, жировая ткань), скелетные мышцы.
Влияние на обмен веществ. По механизму действия глюкокортикоиды относятся к гормонам, проникающим в клетку, и, следовательно, влияют на экспрессию генов и синтез определенных белков. Само название «глюкокортикоиды» обусловлено способностью стимулировать образование глюкозы.
1. Обмен углеводов. Глюкокортикоиды способствуют повышению концентрации глюкозы в крови по следующим механизмам: 1) стимулируют высвобождение аминокислот – субстратов глюконеогенеза – из периферических тканей (мышечная, лимфоидная) через активацию катаболических процессов; 2) увеличивают скорость глюконеогенеза в печени путем повышения количества (и активности) ключевых ферментов глюконеогенеза (пируваткарбоксилазы, фосфоенолпируваткарбоксикиназы, фруктозо-1,6-бисфосфатазы и глюкозо-6-фосфатазы); 3) глюкокортикоиды тормозят потребление и использование глюкозы во внепеченочных тканях (мышцы). В результате увеличивается уровень глюкозы в плазме крови. У здоровых людей это влияние уравновешивается инсулином, оказывающим противоположный эффект; 4) Глюкокортикоиды увеличивают запасы гликогена в печени путем увеличения активности гликогенсинтазы.
Регуляторное действие глюкокортикоидов, обеспечивающее длительное повышение концентрации глюкозы в крови, необходимо для питания клеток мозга при хроническом стрессе.
2. Обмен липидов. Глюкокортикоиды в мозговом веществе надпочечников индуцируют синтез метилтрансферазы, которая стимулирует образование адреналина из норадреналина и таким образом косвенно активирует липолиз в жировой ткани и увеличивает концентрацию свободных жирных кислот в крови. Избыточное количество глюкокортикоидов стимулирует липолиз в одних частях тела (конечности) и липогенез – в других (лицо и туловище).
3. Обмен белков. Глюкокортикоиды в целом оказывают анаболическое действие на обмен белков в печени и почках и катаболическое – в других органах (мышцы, лимфоидная ткань, жировая ткань, кожа и кости). Механизм анаболического действия обусловлен активацией специфических генов и соответствующим возрастанием скорости биосинтеза специфических белков. Блокада синтеза белка в лимфоидной ткани и активный протеолиз в ней увеличивают фонд аминокислот, которые поступают в кровь. В печени и почках эти аминокислоты используются для биосинтеза белка.
4. Иммунная система. Глюкокортикоиды в высокой концентрации тормозят иммунологический ответ организма-хозяина. Они вызывают гибель лимфоцитов и инволюцию лимфоидной ткани, что полезно для подавления реакции отторжения при пересадке тканей.
5. Глюкокортикоиды обладают противовоспалительным действием. Механизм противовоспалительного действия обусловлен способностью ингибировать синтез простагландинов через ингибирование фермента фосфолипазы А2. Кроме того, глюкокортикоиды способствуют поступлению в костный мозг, лимфоидную ткань и селезенку лимфоцитов, моноцитов и эозинофилов и выходу полиморфноядерных лейкоцитов из костного мозга. Они тормозят накопление лейкоцитов в участках воспаления, но стимулируют высвобождение из лейкоцитов веществ, участвующих в воспалительных реакциях (кининов, плазминоген-активирующего фактора, простагландинов и гистамина). Сочетание указанных эффектов приводит к плохому заживлению ран, повышенной чувствительности к инфекции и снижению воспалительного ответа, что обычно наблюдается у людей с избытком глюкокортикоидов. В костной ткани глюкокортикоиды тормозят деление клеток и синтез коллагена. Продолжительное действие этих гормонов приводит к остеопорозу.
При длительном стрессе в крови обнаруживается повышенная концентрацию глюкозы, аминокислот, жирных кислот, кетоновых тел, в моче – глюкозурия, кетонурия, аминоацидурия. Это состояние называется стероидным диабетом.

