




Одним из главных материалистических достижений дарвинизма явился отказ от признания ламарковского внутреннего стремления к совершенству как одной из причин эволюции. (В гл. 1 мы говорили об отношении Ч. Дарвина к прогрессу, который считал, что нет направленного прогресса, нет повышения организации в эволюции.) Однако в целом повышение организации живых структур в макроэволюции явно прослеживается, о чем говорит развитие жизни от пробионта до высшего животного или растения. Отсюда и возникает неудовлетворенность дарвинизмом, поскольку повышение приспособленности, как правило, связано с усложнением структур, а в дарвинизме этого не объясняется. В то же время хорошо известны и широко распространены случаи дегенерации форм, потери ими тех сложных надстроек, которые приобретались в процессе длительного действия отбора на выживание наиболее приспособленных. Самоорганизация жизни, лежащая в основе представлений дарвинизма о движущих силах развития, никак не может объяснить явление дегенерации форм организмов в эволюции. (Зачем нужно упрощение на общем пути к самоусложнению?) Отсюда—прямой путь к агностицизму и к признанию непредсказуемости действия естественного отбора. Получается, что отбор делает, что хочет («выступает в роли господа бога» — по А. А. Любищеву), то усложняя, то, наоборот, резко упрощая структуру. Остается только post factum фиксировать все то, что он «напридумывал» в результате «мгновенного принятия решения», т. е. перейти на описательный путь и попытаться классифицировать имеющееся. Это и было тщательно проделано дарвинистами при разработке учения о разных формах прогресса. Наиболее обстоятельно учение о прогрессе освещено в трудах А. Н. Северцова [1922,1967].
Он был вынужден признавать, что общая дегенерация является полной противоположностью морфофизиологического прогресса, или ароморфоза. Действительно, у многих дегенерировавших форм целиком редуцируются органы движения, т. е. вся мускульная система, а также скелет конечностей, редуцируется центральная нервная система со всеми нервами и органами чувств и органы активного питания и, наоборот, прогрессивно развивается половая система [Северцов, 1967]. Очень существенным замечанием А. Н. Северцова является то, что он говорит о понижении и самой «интенсивности проявления жизнедеятельности при дегенерации вместе с понижением сложности организации» [Там же, т. 138]. Таким образом, дегенерация и ароморфоз полностью противопоставляются по направлению изменений как структуры, так и функции. Но тем не менее оба типа изменений могут приводить к процветанию вида, а это и означает, что их следует включить в эволюционный процесс как равноценные направления, приводящие к биологическому прогрессу (вместе с идиоадаптациями и ценогенезами). Что же, на этом пути остается еще более абстрагироваться, признать единство противоположностей и перестать искать естественно-научное, в данном случае биологическое, объяснение различия форм прогрессивного развития. По-видимому, в пределах только субстратного подхода не удается сделать большего, чем констатация имеющегося. Об этом говорил и А. Н. Северцов [1967], ограничивая рассмотрение эволюции животных тем, «как она происходит в действительности» и не рассматривая, «почему происходит именно то, а не другое изменение» (с. 125).

Рис. 18. Схема возможных изменений в системе «структура — функция» организма (I) и разрешенные (с позиций С + Э подхода) изменения структуры и функции в эволюции: А — направление дегенерации, Б — идиоадаптации и ценогенезы, В — ароморфозы (II). Заштрихована запрещенная зона.
Попробуем, в дополнение к субстратному, применить энергетический подход, т. е. использовать С + Э концепцию. Само собой разумеется, что всякое эволюционное изменение — это изменение и структуры, и функции организма. Схематически все возможные сочетания таких изменений представим в фазовой плоскости «структура—функция» (рис. 18). Стрелка 3 означает, что усложнение структуры связано с усложнением функции, в нашем случае — с увеличением потока энергии, перерабатываемого данным типом биологической структуры. Это как раз и соответствует прогрессивному изменению с позиций структурного подхода. Направлениями от стрелки 2 до стрелки 4 с разными углами наклона и будут иллюстрироваться прежде всего ароморфозы. Заметим, что сюда же следует отнести и идиоадаптации, так как энергетический подход требует хоть и небольшого, но все же усиления функции. Начиная со стрелки 4, включая 5, 6, 7 и 8, мы имеем явное ухудшение функции, что запрещается энергетическими принципами и потому не должно реализовываться. «Самыми запрещенными» являются направления, близкие к стрелке 5, т. е. усложнение структуры и ослабление функции.
Направления, примыкающие к стрелке 1, и будут характеризовать явление дегенерации, так как структура явно упрощается. Но при этом энергетический подход требует усиления функции и не разрешает ее ослабления (по крайней мере в удельном выражении). Термин «общее понижение жизнедеятельности» требует здесь некоторого уточнения. Если организм, популяция вынуждены переходить в новые, часто более простые условия существования (пещеры, большая глубина воды, отсутствие света, глубокие илы и т. д.), то по сравнению со старым местообитанием его функционирование ухудшается. И по-видимому, только упрощение структуры, избавление от ненужных на новом месте старых приспособлений позволяет сохранить и одновременно интенсифицировать функцию, т. е. повысить жизнедеятельность. Таким образом, учет энергетического подхода резко сужает разрешенные варианты развития в системе «структура — функция». Кстати, снимаются и противоположные направления, т. е. тот произвол, что был в рамках структурного подхода (см. рис. 18, II).
Очень существенно то обстоятельство, что эволюция «вниз» по функции не разрешается, если учитывать энергетические принципы. Поэтому говорить о том, что «микроорганизмы эволюционируют в стороны и вниз, но не вверх», с позиций С + Э подхода некорректно, ибо функция улучшается — такая трактовка допустима только с позиций субстратного подхода. Современные микроорганизмы отличаются от древних форм не только разнообразием структур (и иногда их серьезным упрощением), но и существенным усложнением функций: сюда входят и использование различных источников энергии и вещества, недоступных высшим формам, и интенсификация функции до уровней, тоже недостижимых для высших организмов. Но, совершенствуя и во многих случаях упрощая структуру, они под действием естественного отбора интенсифицируют функцию, т. е. эволюционируют «вверх». Как мы обсуждали в предыдущем разделе, питание растворенными веществами делает невыгодным увеличение размеров именно с позиций энергетики. Подчиняясь этому, прокариотные формы выполняют свою миссию в круговороте, «не беспокоясь» об усложнении структур, а наоборот, в соответствии с энергетическими принципами, «заботясь» об освобождении от любой структурной избыточности. Именно на микроорганизмах может быть ярче всего продемонстрирована способность необычайно быстро избавляться от лишних структур.
Как замечает Э. Брода в книге «Эволюция биоэнергетических процессов» [М., 1978, с. 61–62]: «Организмы имеют тенденцию утрачивать ненужные гены и ферменты. Это явление играет огромную роль в эволюции, и особенно в эволюции микробов...
...Автотрофы имеют тенденцию превращаться обратно в гетеротрофы, если дать им подходящие питательные вещества. И вообще, готовность к утратам — это свойство клеток, которое было хорошо изучено на простых системах». Одним из наиболее изученных и показательных фактов «гибкости» приспособления на уровне микробных популяций является смена прототрофных форм в популяции на ауксотрофные. Обычно на обедненных средах прототрофные варианты, способные производить для себя все необходимые метаболиты, быстро вытесняют ауксотрофные формы, требующие для своего развития дополнительно какого-либо определенного соединения. Если же это соединение добавлено в среду, то ауксотроф получает преимущество и способен расти быстрее, так как более экономен и не производит этого соединения в отличие от прототрофов. Этот пример с победой «дефективного» ауксотрофа при развитии на обогащенной среде может служить хорошей моделью явления дегенерации формы при переходе к паразитизму. (Дефективность, т. е. упрощение структуры ауксотрофа, может быть связана с потерей гена (делецией) в ДНК клетки, т. е. иметь генетическую природу.)
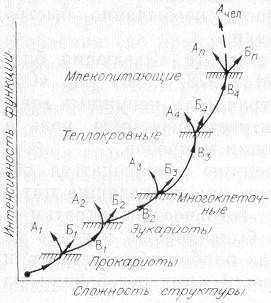
Рис. 19. Общая схема относительного изменения структуры и функции организмов в макроэволюции.
1 — прокариоты; 2 — эукариоты; 3 — многоклеточные; 4 — теплокровные; n — млекопитающие. Буквенные обозначения те же, что и на рис. 18.
Покажем схему усложнения структуры и функции организмов, начиная с пробионтной клетки и кончая млекопитающими, в том числе и человеком (рис. 19). В этом ряду несомненно усложнялась структура, но закреплялись только те варианты, которые приводили к существенному усилению функции, как того требует С + Э подход. Практически на каждом этапе, связанном с крупным ароморфозом, помимо усложнения структуры наблюдалась и дегенерация формы (структуры). На рисунке это отмечено стрелками А, загнутыми влево и вверх. Заметим, что к стрелкам А, указывающим на дегенерацию структуры млекопитающих, по-видимому, должна примыкать и стрелка Ачел, соответствующая человеку. Действительно, что касается структуры, то мы в эволюции немало потеряли: шерсть, спасающую от холода; острые клыки хищника; возможно, и хвосты, помогающие предкам лазать по деревьям. Но энергетика человека как вида возросла в десятки раз (см. гл. 9).
Общая тенденция увеличения угла наклона кривой отражает тот факт, что с повышением организации функция растет быстрее, и это во многом связано с цефализацией структуры. Следовательно, для более точных представлений о темпах эволюции необходимо привлекать и информационный подход. К этому же можно добавить, что возможность какого-либо ароморфоза у млекопитающих, связанного с сильным изменением структуры, мало вероятна. Эволюция из биологической трансформировалась в технологическую, особенно если иметь в виду энергетику человечества. Став глобальной силой, человечество способно вмешиваться в ход эволюции, переведя ее на направленный отбор, селекцию и регулируемую перестройку генотипов.
Хотелось бы обратить внимание на всеобщность и распространенность явления конвергенции, во многих случаях тесно связанного с явлением регресса формы. Конечно же, общими физическими законами определяются особенности и конкретные проявления конвергентности признаков на различных уровнях организации. Среди этих законов хорошо прослеживается действие обоих энергетических принципов. Как правило, конвергентное развитие приводит к наилучшему структурному варианту, позволяющему организму как можно больше изымать энергии (пищи) из окружения, при этом минимизируя собственные траты энергии на перемещение и ориентацию в пространстве и его использование.
Именно сходство условий существования определяет сходство форм и структур различных, а иногда и очень отдаленных таксонов. Это значит, что сходные признаки контролируются очень разными генами или генными комплексами. А это, в свою очередь, значит, что концепция: «Гены хотят жить и размножаться» — явно не состоятельна. Структура оказывается вторичной, подчиняясь функции, которая определяется местом организма (популяции, вида) в круговороте. Это заявление не имеет оттенка телеологичности, ибо с позиций С + Э подхода хорошо известно, что движущая сила развития — поток энергии — имеет физическую природу (а значит, и не имеет каких-либо целей).
Для оценки количественных аспектов изменения формы во времени требуется привлечение информационного подхода, т.е. использование С+Э+И концепции. При этом субстратный подход определит преемственность и модификации структур, энергетический — одну из главных функций, а информационный — скорости перестроек и развития. Можно надеяться, что на этом пути мы сумеем хотя бы в целом вычислять и прогнозировать эволюцию формы и функции и не только на малых отрезках времени.

