




Кафедра зоологии беспозвоночных
Реферат
по дисциплине «Зоология беспозвоночных»
на тему «Возникновение и развитие целомической полости беспозвоночных
животных»
Выполнила
студент группы № 01205
_________ Е.В. Вех
Проверил
доцент
_________ Р. Т-о. Багиров
____________________
зачтено/не зачтено
Томск 2012
Содержание
Введение.....................................................................................................................
Надтип Первичноротые (Protostomia)................................................................... 3
Тип Кольчатые черви (Annelida)......................................................................... 3
Тип Моллюски (Mollusca).................................................................................... 6
Тип Членистоногие (Arthropoda)......................................................................... 7
Тип Онихофоры (Onychophora).......................................................................... 9
Тип Мшанки (Bryozoa)..................................................................................... 10
Тип Плеченогие (Brachiopoda).......................................................................... 10
Развитие целомической полости у трохофорных животных........................... 12
Надтип Вторичноротые (Deuterostomia).............................................................. 14
Тип Иглокожие (Echinodermata)........................................................................ 14
Тип Погонофоры (Pogonophora)....................................................................... 16
Заключение................................................................................................................
Список использованной литературы........................................................................
Введение
Цель данной работы заключается в том, чтобы раскрыть понятие целомической полости (целома), описать ее структуру и функции. Соответственно, главная задача – это изучить внутреннее строение некоторых систематических групп беспозвоночных, чтобы получить представление об эволюции развития целома.
Целом – это вторичная полость тела многоклеточных животных; отличается от первичной полости тела наличием собственной эпителиальной ткани и несколькими другими особенностями, которые будут рассмотрены ниже. Беспозвоночные животные, имеющие в своем строении эту многофункциональную вторичную полость тела, выделяются в особую группу целомических животных (Coelomata). В эту группу входят такие типы, как Кольчатые черви (Annelida), Моллюски (Mollusca), Членистоногие (Arthropoda), Онихофоры (Onychophora), Мшанки (Bryozoa), Плеченогие (Brachiopoda), Иглокожие (Echinodermata), Погонофоры (Pogonophora), Щетинкочелюстные (Chaetognatha), Полухордовые (Hemichordata), Хордовые (Chordata). Далее рассмотрим характерные черты и строение некоторых из них.
Надтип Первичноротые (Protostomia)
Тип Кольчатые черви (Annelida)
Тело представителей этого типа покрыто однослойным эпителием (местами мерцательным), который выделяет на своей поверхности кутикулу. Под эпителием находится хорошо развитый мускульный мешок, состоящий из наружных кольцевых и внутренних продольных мышц. Под кожно-мускульным мешком находится обширная полость тела – вторичная, или целом (отсутствует только в головном и заднем отделах тела). Целом каждого сегмента не сплошной, он поделен двухслойной клеточной перегородкой на правую и левую половины. Перегородка – это складка перитонеального эпителия по срединной спинной линии, спускается книзу, слои ее охватывают с 2 сторон кишечник, под которым вновь соединяются и, достигая брюшной стенки тела, переходят опять в перитонеальный эпителий. Продольная перегородка – брызжейка, или мезентерий, подвешивает кишечник к стенкам тела. На границе между соседними секментами тела кольчецов перитонеальный эпителий образует двухслойную перегородку – септу (диссепимент), пресекающий полость тела. Септами целом делится на известное число поперечных участков, обычно соответствующих числу сегментов тела (но внекотрых отделах септы могут быть редуцированы). Целом заполнен водянистой жидкостью, в которой плавают амебоидные клетки. У некоторых аннелид (пиявки) целом в большинстве случаев вытеснен паренхимой, заполняющей пространство между органами, а целом представлен узкими каналами (лакунарными сосудами) (Островерхова, 2005). Подробное внутреннее строение кольчецов (в т.ч. и целома) показано на рис. 1.

Рис. 1. Поперечный разрез из средней части тела Lumbricus terrestris (Догель, 1981): 1 - кутикула; 2 - эпидермис; 3 - слой кольцевой мускулатуры; 4 - слой продольной мускулатуры; 5 - целомический эпителий; 6 - метанефридий; 7 - щетинка; 8 - мезентерий; 9 - брюшной сосуд; 10 - субневральный сосуд; 11 - брюшная нервная цепочка; 12 - хлорагогенные клетки; 13 - полость кишки; 14 - сосудистый плексус; 15 - тифлозоль; 16 - сосуд тифлозоля; 17- спинной сосуд.
Функции целома:
1. Опорная: целомическая жидкость, близкая по составу к воде, обеспечивает несжимаемость тела; при сокращении кожно-мускульного мешка стенки тела давят на целомическую жидкость и все тело приобретает значительную упругость (ригидность);
2. Распределительная: питательные вещества из кишечника распределяются по организму;
3. Выделительная: в целом поступают и накапливаются жидкие и твердые продукты обмена веществ (экскременты);
4. Половая: в целоме происходит созревание половых клеток (сперматозоидов и яйцеклеток) (Островерхова, 2005).
Интересно, что у класса Первичных кольчецов (Archiannelida) преобладает первичная полость тела, а несегментированный целом (спинной и брюшной) развивается только в период полового размножения. И после того, как в нем разовьются гонады, он редуцируется (Догель, 1981).
У некоторых представителей класса Пиявок (Hirudinea) вторичная полость тела (целом) заменяется системой лакунарных каналов. Они имеют толстые мускулистые стенки, сокращение которых обуславливает движение целомической жидкости по телу. Лакуны могут выполнять часть функций кровеносной сиситемы, жидкость внутри них принимает характер крови, окрашена в красный цвет, содержит гемоглобин и амебоидные клетки. Стенки части лакун покрыты скоплениями зернистых клеток, в которых накапливаются нерастворимые продукты обмена веществ, содержимое которых затем выводится через целом и нефридии из организма (Островерхова, 2005).
Животные класса Эхиурид (Echiurida) обладают обширным, занимающим почти всю внутренность тела целомом. Он сообщается с задней кишкой через 2 анальных мешка, мелкие воронки которого открываются в целом.
Целом у представителей класса Сипункулид (Sipunkulida) так же выстлан перитонеальным эпителием, находится внутри кожно-мускульного мешка и сообщается с брюшным нервным стволом. Целом разделен поперечной перегородкой на 2 участка. Жидкость целома розовая, мутная от многочисленных клеточных включений. Разнообразие плавающих в ней клеток чрезвычайно велико: это амебоидные клетки с псевдоподиями, округлые сплющенные розовые клетки (из-за присутствующего в них железосодержащего гемэритрина). Таким образом, эти клетки можно приравнять к красным кровяным тельцам позвоночных, а сама полостная жидкость, несомненно, играет роль крови. Также своеобразными включениями являются урночки, формирующиеся за счет стенок целома и принимающие участие в выделении экскретов. Они представляют собой двуклеточные тельца, одна клетка в виде прозрачного купола, другая прижата к основанию купола и усажена ресничками, благодаря которым урночки быстро плавают в полостной жидкости. Накапливая продукты обмена, они склеиваются и образуют так называемые желтые тела, выводимые из организма через нефридии. Кроме урночек в целоме плавают диски – овальные пластинки, сами по себе неподвижные, но при повреждении кишки приклеивающиеся к месту поражения с помощью подвижных амебоидных клеток. В полостной жидкости находятся также созревающие половые клетки (Островерхова, 2005).
Целом имеется не у всех кольчатых червей и в разных группах имеет разные онтогенетические источники. Так, целом может развиваться не только телобластически, т.е. иметь мезодермальное происхождение из клеток-телобластов, но и энтодермальное, развиваясь энтероцельно из крыши первичной кишки. Кроме того, целом у кольчатых червей может возникать из схизоцеля, или мускульного зачатка - миоцеля, или из половых желез. В общем целом кольчецов может быть не развит (примитивные Archiaimelida и Polychaeta), может быть целостным (Sipunculida, Echiurida, и некоторые Archiannelida), но у большинства - метамерный с перегородками (Островерхова, 2005).
Тип Моллюски (Mollusca)
Целом моллюсков неметамерный, остаточный, он не связан с внутренними органами так сильно, как у кольчатых червей, включает в себя лишь околосердечную сумку – перикардий и полость гонад. Он закладывается в эмбриональном развитии, и затем постепенно редуцируется, а промежутки между органами заполняются вместо жидкости соединительной тканью. Целомодукты, открывающиеся в перикардий, выполняют функцию почек, а те, что открываются в полость гонад, служат половыми протоками (Догель, 1981).
У представителей раковинных моллюсков класса Моноплакофоры (Monoplacophora) кроме перикардиальных целомов имеются очень обширные парные дорсальные целомы (схематичное строение которых изображено на рис. 2), сообщающиеся с внешней средой с помощью целомодуктов и так же выполняющие функции почек. Эти почки связаны протоками с яичниками и семенниками, помогая выводу половых продуктов наружу (Догель, 1981).

Рис. 2. Схема поперечного разреза Neopilina (Догель, 1981): 1 - аорта; 2 - дорсальный целом; 3 - раковина; 4 - почка; 5 - плевровисцеральный ствол; 6 - жабра; 7 - нога; 8 - педальный ствол; 9 - половая железа; 10 - кровеносная лакуна; 11 –кишка.
Тип Членистоногие (Arthropoda)
Этот тип интересен тем, что отдельного целома у них не существует, зато имеется миксоцель, образовавшаяся за счет слияния первичной полости тела (бластоцеля) и вторичной (целома). Полостная жидкость, наполняющая миксоцель, представляет собой гемолимфу, или кровь. Миксоцель разделяется диафрагмами на синусы: перикардиальный, висцеральный, периневральный. В перикардиальном синусе расположено сердце, в периневральном – брюшная нервная цепочка, а в висцеральном – остальные внутренние органы. Остатки целома, так же, как и у моллюсков сохраняются в почках и гонадах.
Видоизмененные целомодукты – это протоки целома, открывающиеся наружу. Они образуются за счет периферического цитобласта, и во многих случаях совместно с мальпигиевыми сосудами принимают на себя выделительную функцию. При этом целомодукты могут совершенно заменять настоящие протонефридии (у моллюсков), или вступать с ними в многообразные отношения (у полихет). Из всех классов членистоногих более постоянно они сохраняются у ракообразных. У них имеются выделительные железы в двух сегментах: в сегменте антенн II (антеннальные) и максилл II (максиллярные). Выделительные железы ракообразных состоят из двух отделов: концевого мешочка (остаток целома) и выводного канала, открывающегося воронкой в мешочек и образующегося разрастанием стенок целома. Наряду с экскреторной функцией целомодукты ракообразных несут и функцию осморегуляции, т.е. поддерживают постоянство солевого состава и количество свободной воды в клетке (Островерхова, 2005).
Подтип Трахейнодышащие (Tracheata)
Рассмотрим Классы Губоногие многоножки (Chilopoda) и Двупарноногие многоножки (Diplopoda). Их выделительная система представлена 1-2 парами мальпигиевых сосудов. Влага из экскретов, поступающих из мальпигиевых сосудов, всасывается в стенку задней кишки и снова поступает в обменные процессы. Кровеносная система незамкнута, и кровь из артерий выливается в лакуны миксоцеля. Из лакун кровь собирается в брюшной венозный сосуд, а из него в околосердечный синус, затем через ости поступает снова в сердце (Шарова, 2004).
Подтип Жабродышащие (Branchiata)
У класса Ракообразных (Crustacea) миксоцель частично выполняет кровеносную функцию, кровь движется из сердца (строение которого сильно различается у разных видов) по нескольким крупным сосудам в полость тела, откуда по системе вен течет в жабры, чтобы насытиться кислородом.
Выделительные органы раков представлены двумя парами видоизмененных целомодуктов - железистых органов. Одна пара открывается у основания антенн (антеннальные железы), другая - у основания второй пары нижних челюстей (максиллярные железы). Строение обеих пар приблизительно одинаково: каждый орган состоит из концевого мешочка и отходящего от него извитого канала с железистыми стенками; канал делает несколько петлеобразных изгибов и затем открывается наружу, иногда образуя перед этим заметное расширение - мочевой пузырек. Установлено, что концевой слепо замкнутый мешочек - это сохранившийся от эмбрионального состояния участок целома, а отверстие, сообщающее пузырек с каналом, - воронку целомодукта, но лишенную мерцательных ресничек. Одновременное присутствие обеих пар выделительных желез встречается. Обычно же одна из них функционирует на личиночных стадиях и потом редуцируется, заменяясь во взрослом состоянии другой. При этом все раки во взрослом состоянии имеют максиллярные железы, кроме высших раков, имеющих антеннальные железы (Островерхова, 2005).
Тип Онихофоры (Onychophora)
У этих беспозвоночных отсутствует хитиновый скелет, имеется кожно-мускульный мешок, под которым находится смешанная полость тела – миксоцель. Формирование миксоцели показано на рис. 3.



Рис. 3. Развитие целома у Peripatus (Догель, 1981): А - поперечный разрез зародыша; целомические мешки подразделены на 3 отдела: спинной, брюшной и боковой; Б, В-то же, превращение спинных отделов в половые железы и образование целомодуктов за счет брюшного и отчасти бокового отделов. Стенка значительной части боковых отделов разрушается, и на их месте образуется смешанная полость тела (миксоцель). 1 - спинной отдел целома; 2 - боковой; 3 - брюшной; 4 - брюшной орган; 5 - кишка; 6 - нервный ствол; 7 - половая железа; 8 - сердце; 9 - концевой пузырек целомодукта
Производные целома представлены полостью гонад и целомодуктами, которые метамерны по строению и выполняют выделительную функцию (Догель, 1981).
Полость тела у класса первичнотрахейных (Protracheata) в общем сходна с таковой членистоногих. У зародышей образуются парные метамерные целомические мешки, которые позднее разрушаются, так что вторичная полость тела смешивается с первичной. Из клеточного материала целомических мешков формируется мускулатура, соединительная ткань и т.д. Сохраняются лишь небольшие участки мешков, превращающихся в пузырьки выделительных органов и дающие стенку гонад. Однако кроме этих остатков целома сохраняется также перитонеальный эпителий на поверхности внутренних органов (кишечника и др.) (Островерхова, 2005).
Тип Мшанки (Bryozoa)
Тело их покрыто однослойным эпителием, кожно- мускульный мешок развит слабо из-за неподвижного образа жизни. Перитонеальный эпителий ограничивает обширную вторичную полость тела, в которой помещаются все внутренности. Целом делится тонкими перегородками на 3 отдела. Передний – небольших размеров и залегает в эпистолеме. Средний - кольцевой канал – окружает глотку и посылает слепые ответвления в щупальца. Задний, наиболее обширный, занимает почти все тело и называется туловищным целомом. У голоторотых мшанок эпистом вместе с целомом редуцируется. Нет кровеносной системы и выделительных органов. У покрыторотых мшанок полости целомов соединяются трубчатыми образованиями, которые рассматриваются как видоизмененные целомодукты. Также у некоторых видов мшанок существует особая целомическая пора, расположенная между щупальцами и ротовым отверстием и служащая для вывода оплодотворенных яиц во внешнюю среду (Островерхова, 2005).
Тип Плеченогие (Brachiopoda)
Тело одето однослойным кожным эпителием, под которым находится тонкий слой соединительной ткани, а затем мерцательный перитонеальный эпителий, выстилающий обширную вторичную полость тела. Сплошной кожно-мускульный мешок отсутствует вследствие развития раковины, которая не позволяет телу сокращаться и изменять форму, но сильно развиты отдельные мышечные пучки, идущие через полость тела от внутренней поверхности одной створки к другой (Шарова, 2004).
Целом плеченогих представлен обширной полостью тела, содержащей кишечник и пересекаемой мышцами раковины. Мезентерий делит целом на правую и левую половины; целом дает выросты, проникающие в толщу мантийных складок, а также узкие каналы, проходящие внутри рук. От этих каналов отходят тонкие веточки в каждое щупальце. В полости тела плеченогих, кроме продольных мезентериев, имеются и неполные поперечные перегородки. Иногда их рассматривают как остатки настоящих диссепиментов, разделяющих целом на три отдела: передний - эпистомальный; средний, посылающий целомические каналы в лофофор и щупальца, и задний - туловищный. Полость тела содержит бесцветную жидкость с рассеянными в ней амебоидными клетками (Островерхова, 2005). Подробное внутреннее строение целома плеченогих показано на рис. 4.
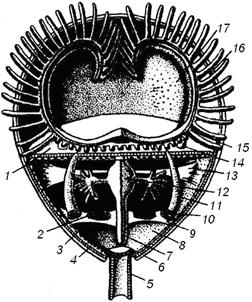
Рис. 4. Схема строения плеченогого (Догель, 1981): 1 - передняя стенка тела; 2 - яичник; 3 - перитонеальный эпителий; 4 - кожный эпителий; 5 - стебелек; 6 - раковина; 7 - средняя кишка; 8 - целом; 9 - задняя складка перитонеального эпителия; 10 - воронка целомодукта; И - целомодукт; 12 - желудок; 13 - передняя складка перитонеального эпителия; 14 - основание руки; 15 - пищевая бороздка между основаниями щупалец; 16- спинная мантийная складка; 17- мантийная полость
Развитие целомической полости у трохофорных животных
Трохофорные, или членистые, животные (Trochozoa) включают типы: Моллюски (Mollusca), Сипункулиды (Sipunculida), Эхиуриды (Echiurida), Кольчатые черви (Annelida), Тихоходки (Tardigrada), Онихофоры (Onychophora), Членистоногие (Arthropoda). Для большинства из них характерна пелагическая личинка - трохофора, обладающая слепым или сквозным кишечником, первичной полостью тела, протонефридиями. В силу своеобразия и преобладающего среди трохозоев числа целомических животных вопрос сместился в сторону выяснения их происхождения.
У трохофорных животных, эволюция которых была направлена по пути развития метамерии, целом в разных группах Trochozoa то приобретал, то терял метамерное строение. В связи с этим в эволюции трохозоев выделяется три основных направления, представленных соответствующими группами, характеризующимися функциональными преобразованиями целома и разной степенью развития метамерии.
Первая группа характеризуется исходным нерасчлененным состоянием целома, не связанным с метамерией тела. Современные Echiurida, Priapulida, вероятно, являются реликтовыми группами таких первично несегментированных Coelomata, причем целом у них возник независимо путем эпителизации стенок схизоцельных (первичных) опорных полостей. Схизоцельный целом у животных появляется раньше, чем развивается истинная метамерия.
Вторая группа представлена в основном аннелидами (Annelida), имеющими разную степень и формы расчленения целома. В ряду трохофорных животных развитие метамерного целома кольчецов больше всего связано с развитием метамерии тела, физиологией, экологией и особенностями способов движения. У полихет, олигохет и пиявок можно наблюдать несколько разных способов движения и соответственно разное строение целома.
Так, развитие многочисленных парных целомических камер у свободноживущих полихет функционально связано с изгибанием их тела в горизонтальной плоскости и сопровождается развитием параподиальных выростов. Преобладание перистальтического способа движения у роющих полихет и олигохет сопряжено с частичной утратой поперечных перегородок полости тела и в ряде случаев параподиальных выростов. У пиявок преобладает изгибание тела в вертикальной плоскости. При таком способе движения парные целомические камеры теряют свое функциональное значение. Опорная функция в этом случае переходит к паренхиме, а гомеостатическая, транспортная и выделительная функции сохраняются за целомической полостью, которая приобретает форму лакунарной системы.
Появление целома у предков современных целомических животных - это ароморфоз, открывший новые пути для адаптивной радиации многообразной группы Coelomata. По поводу источников происхождения целома существует несколько теорий: схизоцельная, миоцельная, гоноцельная и энтероцельная. Основные различия между этими теориями (гипотезами) заключаются в том, какая из многих функций целома принимается за первичную и какой процесс формирования целома принимается за первичный (Островерхова, 2005).
Схизоцельная теория рассматривает целом как гомологичное образование первичной полости тела Nemathelminthes, представляющее простое ее усовершенствование. Эта теория не объясняет, почему при эмбриональном развитии у многих животных целом формируется за счет стенок кишечника.
По миоцелъной теории констатируется образование целома как полости в мускульном зачатке животного, которая заполнилась жидкостью и выполняла опорную функцию для окружающей мускулатуры. Другие функции целома эта теория не объясняет (Догель, 1981).
По гоноцельной теории целом производится из половых желез низших червей. Согласно этой теории, полость каждого целомического мешка соответствует полости гонады турбеллярии или не-мертины, перитонеальный эпителий происходит из стенки половой железы, а целомодукты - из половых протоков. В этой теории много натяжек.
Сторонники энтероцелъной теориисчитают, что целом берет начало от гастроваскулярной системы кишечнополостных (Coelenterata) и гребневиков (Ctenophora), которые рассматриваются как отдаленные предки целомических животных. Так как у высших кишечнополостных под энтодермой гастроваскулярных каналов развиваются гонады, то энтероцельная теория хорошо объясняет половую функцию целома. Она, по существу, включает в себя и гоноцельную теорию (Островерхова, 2005).
Надтип Вторичноротые (Deuterostomia)
Тип Иглокожие (Echinodermata)
Целомическая полость данных беспозвоночных эволюционировала до так называемой амбулакральной системы, являющейся одной из наиболее оригинальных черт их строения. Амбулакральная (водно-сосудистая) система представляет собой обособленную часть вторичной полости тела и содержит жидкость, близкую по составу к морской воде, с примесью белка и присутствием амебовидных клеток (Шарова, 2004).
Образование амбулакральной системы начинается с изменения целомических мешков. Каждый из них перешнуровывается дважды, давая справа и слева от кишечника уже три пары целомических мешков, что свидетельствует о 3-сегментном (олигомерном) строении личинки. Оба задних (третьих) мешка преобразуются в целом взрослого животного. 1-й и 2-й (правые) - чаще недоразвиты; основные изменения претерпевают 1-й и 2-й (левые) целомические мешки, между которыми остается связь в виде канальца, превращающегося в дальнейшем в каменистый канал. При этом первый целомический мешок получает сообщение с наружной средой особым канальцем с отверстием - будущим мадрепоритом; сам же первый целомический мешок входит позже в состав осевого органа взрослого иглокожего. По мере метаморфоза средний левый целомический мешок, называемый теперь гидроцель, т.е. зачаток всей амбулакральной системы, вытягивается и охватывает переднюю кишку личинки сначала в виде подковы с пятью слепыми выростами, а затем подкова смыкается в околоротовое кольцо; слепые выросты вытягиваются и дают начало радиальным амбулакральным каналам. Следовательно, всю амбулакральную систему можно толковать как видоизмененный целомический мешок, а каменистый канал - как целомодукт этого мешка, изменивший свою первоначальную функцию (Островерхова, 2005). На рис. 5 показано строение амбулакральной системы иглокожих на примере морского ежа.

Рис. 5. Общая анатомия морского ежа (Догель, 1981): 1 - глазок; 2 – аборальное кровеносное кольцо; 3 – анальное отверстие; 4 – мадрепоровая пластинка; 5 – половая железа; 7- кишечник; 8 - каменистый канал; 9 – осевой орган; 10 – жабра; 11 – скелет аристотелева фонаря; 12 – зуб; 13 – рот; 14 – оральная эктоневральная система; 15 – кишечный кровеносный сосуд; 16 – полиевы пузыри; 17 – кольцеворй канал амбулакральной системы; 18 – оральное кровеносное кольцо; 19 - радиальный амбулакральный сосуд; 20 – радиальный кровеносный сосуд; 21 – радиальный канал перигемальной системы; 22 – ампулы амбулакральных ножек; 23 – амбулакральные ножки; 24 – радиальный нервный ствол
Еще одной особенностью иглокожих является наличие перигемальной (псевдогемальной) системы. Ее строение таково: от общего целома обособляется участок, продолжающийся в лучи и образующий перигемапьную систему каналов. Она образует околоротовое кольцо, залегающее между нервным и амбулакральным кольцами. От кольца отходят радиальные перигемальные каналы, каждый из которых поделен надвое продольной вертикальной перегородкой. Перигемальная система содержит ту же жидкость, что и целом. Перигемальная система сопровождает нервную систему и, вероятно, служит специально для питания нервных тканей и для предохранения их от сдавливания. То есть нервная система подстилается не плотными частями, а омывается перигемалыюй жидкостью.
Амбулакральная система хорошо развита у таких классов, как Морские звезды (Asteroidea) и Морские ежи (Echinoidea). Она начинается на верхней (аборальной) стороне продырявленной пластинкой – мадрепоритом, у некоторых иглокожих вместо него существует только одно небольшое отверстие - гидропора. Поры мадрепорита ведут в небольшую полость, от которой начинается каменистый канал, имеющий в стенках известковые пластинки (отсюда название). Каменистый канал впадает в кольцевой канал, окружающий глотку. От кольцевого канала по радиусам отходят пять боковых, или радиальных, каналов. Между ними расположено пять мешковидных выростов - полиевых пузырей, содержащих запас жидкости. От каждого радиального канала отходят в стороны попарно тонкие трубочки - амбулакральные ножки, каждая из которых заканчивается остроконечно или подошвой. Стенки ножек растяжимы и мускулисты. Амбулакральные ножки связаны с пузырьками-ампулами и выполняют разнообразные функции: газообмен, осязание, движение. Каждый радиальный канал заканчивается чувствительным концевым щупальцем. Радиальные каналы с амбулакральными ножками и связанные с ними скелетные элементы составляют комплексы, называемые иногда амбулакрами, или амбулакральными радиусами. Между амбулакрами расположены так называемые интеррадиусы - интерамбулакры (Островерхова, 2005).
Тип Погонофоры (Pogonophora)
Стенка тела погонофор состоит из однослойного, богатого одноклеточными железами кожного эпителия, который покрыт почти сплошь кутикулой, под ним располагаются кожно-мускульный мешок и перитонеальный эпителий, ограничивающий вторичную полость тела. Все тело делится на четыре первичных сегмента, содержащих каждый свой отдел целома. Кишечник полностью отсутствует, для собирания из воды пищевых частиц и для всасывания питательных веществ служат щупальца, развитые на первом сегменте, а целомодукты первого сегмента служат органами выделения. Третий сегмент очень длинный, на его протяжении многие органы, в том числе участки целома, приобретают более или менее правильное метамерное расположение, его целомодукты также выполняют функцию выведения половых продуктов. Четвертый сегмент отличается правильной вторичной сегментацией (Догель, 1981), как показано на рис. 6.
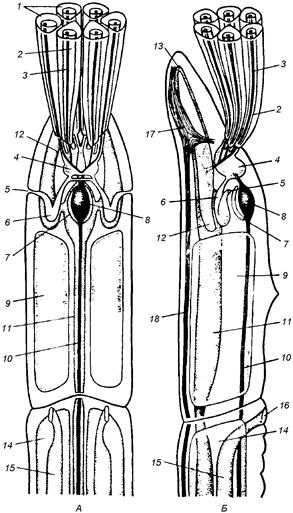
Рис. 6. Схема строения Pogonophora (Догель, 1981): А - передний участок тела самца со спинной стороны; Б- то же с левой стороны: 1- щупальца; 2- целомический канал щупальца; 3 - приносящий и уносящий сосуды щупальца; 4 - целом первого сегмента; 5 - наружное отверстие целомодукта; 6- целомодукт первого сегмента; 7-перикардий; 8-сердце; 9-целом второго сегмента; 10 -спинной сосуд; 11- брюшной сосуд; 12 - боковой головной сосуд: 13 - срединный головной сосуд; 14 - целом третьего сегмента; 15 - семяпровод; 16 - половое отверстие; 17- мозг; 18-брюшной нервный ствол
Кроме этой сегментации (первичной), у погонофор существует и зачаточное вторичное расчленение целома, а именно: от общего целома в передней части туловища отделяются небольшие участки, находящиеся в метамерных брюшных прикрепительных папиллах; в заднем, щетинконосном участке туловища, образуются поперечные перегородки, врастающие от стенок тела в уже существующий здесь общий целом, который разделяется благодаря этому на ряд метамерных камер (Островерхова, 2005).
Заключение
В процессе работы было изучены следующие группы целомических животных (Coelomata): Кольчатые черви (Annelida), Моллюски (Mollusca), Членистоногие (Arthropoda), Онихофоры (Onychophora), Мшанки (Bryozoa), Плеченогие (Brachiopoda), Иглокожие (Echinodermata) и Погонофоры (Pogonophora). Изучив их внутреннее строение, я могу сказать, что целом является очень важной частью их организмов. У беспозвоночных еще не полностью развиты кровеносная, выделительная и половая системы, зато большинство их функций выполняет целом. Находящаяся внутри него жидкость активно участвует в процессе обмене веществ, помогает поддерживать упругость (ригидность) тела.
У разных типов животных целом принимает неодинаковую форму: у членистоногих он преобразуется в миксоцель, у иглокожих развивается в амбулакральную систему, у моллюсков же почти полностью редуцирован. Однако, в каком бы виде целом ни существовал в теле животных, он является прекрасным приспособлением для адаптации беспозвоночных в природе. И в результате можно сделать вывод, что целомические животные действительно находятся на более высокой ступени эволюции, чем первичнополостные животные.
Список использованной литературы:
1. Догель В.А. Зоология беспозвоночных: Учебник для университетов/Под ред.проф. Полянского Ю.И. – 7-е изд. – М: Высшая школа, 1981. – 606 с.
2. Островерхова Г.П. Зоология беспозвоночных: Учебник для университетов. – Томск: Изд-во Том. ун-та, 2005. – 660 с.
3. Шарова И.Х. Зоология беспозвоночных: Учеб. для студ. выс. учеб. заведений. – М: Гуманит. изд. центр ВЛАДОС, 2003. – 592 с.

