




До складу комісії включаються: керівник чи представник служби охорони праці; керівник структурного підрозділу, на якому працює потерпілий; представник профспілкової організації або уповноважений трудового колективу з питань охорони праці, якщо потерпілий не є членом профспілки.
Керівник робіт, який безпосередньо відповідає за охорону праці на місці, де стався нещасний випадок, до складу комісії не включається.
При нещасному випадку з можливою інвалідністю до складу комісії включається також представник Фонду.
Потерпілий або його довірена особа має право брати участь у розслідуванні нещасного випадку.
Смотрите www.beautyline.com.ua лазерное омоложение.
Комісія з розслідування протягом трьох днів зобов'язана:
- обстежити місце нещасного випадку, опитати свідків і причетних осіб, і, по можливості, одержати пояснення потерпілого;
- визначити відповідність умов і безпеки праці вимогам нормативно-правових актів з охорони праці;
- з'ясувати обставини й причини нещасного випадку, визначити зв'язок цього випадку з виробництвом;
- визначити осіб, які порушили вимоги охорони праці, а також розробити заходи щодо запобігання подібних випадків;
- скласти акт розслідування за формою Н-5 у двох примірниках, а також акт Н-1 або НТ про випадок в шести примірниках і передати його на затвердження роботодавцеві.
До першого примірника акта Н-5 (далі – акт розслідування нещасного випадку) додаються акт Н-1 або НТ, пояснення свідків, потерпілих, витяг з експлуатаційної документації, схеми, фотографії та інші документи, що характеризують стан робочого місця, а також, у разі потреби, медичний висновок про наявність в організмі потерпілого алкоголю, отруйних або наркотичних речовин.
Статевий процес у рослин
У більшості еукаріотів (нижчих і вищих рослин) в життєвому циклі обов’язковим є статеве розмноження, оскільки воно має ряд.значних переваг над іншими способами розмножень, а саме: 1) більш високий коефіцієнт розмноження, тобто утворюється більша кількість зачатків нових особин; 2) більша можливість розселення на значно дальші відстані, а відповідно й заселення значних територій; 3) імовірність появи під дією різних умов навколишнього середовища нового матеріалу для природного добору. Статеве розмноження забезпечує повне відновлення організму,
оскільки життя кожної нової особини починається спочатку в новій якості і всі. вікові зміни батьків потомству не передаються. Крім того, статеве розмноження забезпечує поєднання більш-менш різних батьківських і материнських спадкових задатків, потомство стає більш різноманітним, з новими комбінаціями батьківських і материнських властивостей, а іноді з зовсім новими ознаками. Таке генетичне більш "різнорідне потомство забезпечене більш широкою амплітудою пристосування до умов навколишнього середовища. Окремі представники його. можуть жити в таких умовах, де їхні батьки загинули б. Отже, з появою статевого розмноження організми набули більш високої конкурентної спроможності щодо пристосованості їх і виживання.
Статевий процес – це злиття (копуляція) двох статевих клітин (гамет) і утворення однієї клітини – зиготи, тобто зменшення кількості клітин. Отже, термін “статевий процес” (злиття гамет і “статеве розмноження” як збільшення, чисельності особин певного виду, що утворилися в результаті статевого процесу, не можна вважати синонімами, оскільки збільшення статевого потомства відбувається, як правило, за рахунок збільшення кількості гамет і відповідно кількості зигот, ними утворених.
Злиття гамет супроводжується не лише злиттям цитоплазми копулюючих клітин, а й обов’язковим є злиття їхніх ядер. Утворене копуляційне ядро матиме подвійний (2n) набір хромосом, його називають диплоїдним (від гр. diploos – подвійний); ядра гамет мають відповідно вдвічі меншу кількість хромосом (n), їх. називають гаплоїдними (від rp. haploos – простий, одинарний). Злиття ядер – найважливіший етап в статевому процесі, його називають заплідненням.
Для рослин характерні різні типи гамет, відповідно розрізняють різні типи статевого процесу. Типи статевого процесу буде описано в тій послідовності, в якій здійснювалось поступове ускладнення еволюційного шляху його становлення.
Найпростішим типом статевого процесу в нижчих рослин є злиття двох клітин, позбавлених твердої оболонки, які є цілим організмом. Наприклад, в одноклітинної зеленої водорості дуналіели сольової (Dunaliella salina) при статевому розмноженні зливаються не гамети, а звичайні одноклітинні індивідууми, які на певному етапі життя поводять себе як гамети. Такий статевий процес називають хологамією. Зовні копулюючі клітини не відрізняються між собою..
Подібний примітивний тип статевого, розмноження – кон’югація. Вона спостерігається в зеленої нитчастої водорості спірогіри (Spirogira), нитки якої складаються з гаплоїдних клітин. Восени, коли звичайно настає час статевого розмноження, дві нитки розміщуються паралельно і клітини, що лежать одна проти одної, утворюють копуляціині відростки, які, збільшуючись, сполучаються, оболонки розслизнюються, формуючи копуляційний канал між цими клітинами. Протопласт клітин стискається і округляється, а потім поступово через канал з однієї клітини переливається до іншої. Клітину, вміст якої переливається до іншої, прийнято вважати чоловічою, а ту, в якій відбувається злиття, – жіночою. Отже, в кон’югації беруть участь звичайні, не спеціалізовані вегетативні кліти-ни багатоклітинної водорості.
Значно частіше в рослин статевому розмноженню передує поділ клітин і утворення в них спеціалізованих статевих клітин – гамет. Залежно від розміру і рухомості гамет розрізняють кілька типів статевих процесів.
Іноді всі гамети, як чоловічі, так і жіночі, рухомі, однакові за формою та розмірами, їх умовно позначають як плюс- і мінусгамети, в статевий процес такого типу називають ізогамним. З ізогамним статевим процесом в більшості водоростей (Chlamydomonas, Ulotrix і ін.) зливаються гамети різних особин, що, називають гетероталізмом (мал. 20).
У деяких водоростей рухомі гамети розрізняють за розміром – це гетерогамний статевий процес. Багатьом нижчим і всім вищим рослинам властивий оогамний статевий процес. Це форма статевого розмноження, в якому бере участь одна гамета – нерухома, позбавлена джгутиків, що має порівняно більший розмір із значним запасом поживних речовин, ії називають яйцеклітиною, або жіночою гаметою. Друга гамета маленька, рухома, озброєна джгутиками, містить досить велике ядро і незначну кількість цитоплазми – це чоловіча гамета, або сперматозоїд (від гр. sperma – насіння, zoon – жива істота). Приваблені хімічними речовинами, виділеними яйцеклітиною, сперматозоїди підпливають до яйцеклітини і, зливаючись з нею, виконують запліднення. Більшість насінних рослин (переважна більшість голонасінних і всі покритонасінні) в процесі еволюції втратили джгутики в чоловічих гаметах. Безджгутикові чоловічі гамети називають сперміями.
Отже, оогамія – еволюційно найвищий тип статевого розмноження рослин, що має ряд значних переваг: 1) велика за розміром яйцеклітина з запасом поживних речовин забезпечує живлення зиготи доти, доки вона не набуде здатності до самостійного живлення; 2) зменшення за розміром чоловічих гамет забезпечило їм здатність рухатись у дуже малій кількості види або навіть за її відсутності в наземних умовах, а велика кількість і рухомість сперматозоїдів (сперміїв) підвищують ефективність статевого процесу; 3) нерухомість яйцеклітини створила передумови виникнення внутрішнього запліднення, а також надійнішого захисту зиготи, що особливо важливо в умовах наземного існування і найбільш поширене серед вищих рослин.
У більшості рослин статеві клітини формуються в особливих органах – гаметангіях. Гаметангії нижчих рослин, як і їхні спорангії, одноклітинні. У примітивних, рослин гамети утворюються в вегетативних клітинах шляхом поділу їхнього вмісту. Кількість гамет в одному гаметангії, а також кількість гаметангіїв, що формуються однією особиною, може бути різною – від однієї до кількох.
В разі оогамії чоловічі й жіночі статеві органи частіше мор-фологічно відрізняються. Гаметангії, в яких утворюються яйцеклітини, називають оогоніями, чоловічі гаметангії – антеридіями.
У вищих рослин гаметангії багатоклітинні. Жіночі статеві органи називають архегоніями, чоловічі, як і в нижчих, – антеридіями. Однак у насінних рослин, які пройшли значний шлях еволюційних змін, у зв’язку з пристосуванням до наземного існування статеві органи зазнали значної редукції (у голонасінних) або зовсім втрачені (в покритонасінних).
3.1. ПОНЯТТЯ ПРО ЦИКЛ ВІДТВОРЕННЯ
Під циклом відтворення розуміють певний відрізок життя виду, що обмежений двома однойменними етапами: від спорофіту до спорофіту, від гаметофіту до гаметофіту, від мейоспори до мейоспори, від зиготи до зиготи і т. д.
Уже в багатьох високоорганізованих водоростей (зелених, бурих, червоних) виявлено чергування поколінь в одному циклі: нестатевого, яке завершується формуванням спор (зооспор), і статевого, яке закінчується утворенням гамет. Перше з них за кінцевим продуктом називають спорофітом, друге – гаметофітом. Ця закономірність є обов’язковою також для всіх вищих рослин:
мохоподібних, плауноподібних, хвощеподібних, папоротеподібних, голонасінних і покритонасінних.
Зазначене чергування поколінь обов’язково супроводжується зміною ядерних фаз – диплоїдною і гаплоїдною. Диплоїдна фаза починається завжди із зиготи. Характеризується вона подвійним набором хромосом в ядрі (при копуляції гамет хромосоми їх-не зливаються, а лише об’єднуються). Закінчується диплоїдна фаза редукційним поділом клітин (мейозом), при якому кількість хромосом зменшується вдвічі.
Редукційний поділ відбувається в рослин залежно від систе-матичного виду в. певний період циклу відтворення. У більшості зелених, різноджгутикових та інших водоростей мейотична ділиться (після періоду спокою) зигота. У зв’язку з цим все їхнє життя, за винятком зиготи, відбувається в гаплоїдному стані – рослини гаплобіонти. У деяких зелених (ульвових, улотрипксових, хетофорових), бурих і червоних водоростей, а також у всіх вищих рослин редукційний поділ (мейоз) відбувається при утворенні спор ста-тевого розмноження, тобто спорофіт відповідає диплонту, а гаметофіт – гаплонту.
Спори, які виникають внаслідок мейозу, називають спорами статевого розмноження, тому вони під час проростання дають початок статевому поколінню – гаметофіту.
Морфологічна будова, потужність розвитку і тривалість життя гаметофіта відносно спорофіта неоднакова в різних відділах рослин. У багатьох зелених водоростей більш розвинутий гаметофіт, у бурих (диктіоти, ектокарпуса), зелених (ульви, хетофорових), більшості червоних водоростей обидва покоління розвинені однаково. Одночасно, у ламінарії як представника бурих водоростей переважає спорофіт.
У всіх вищих рослин, за винятком мохів, домінуючим є спорофіт, гаметофіт же представлений або невеликим заростком (плауни, хвощі, папороті), або досить редукованим настільки, що розпізнати його можна лише при детальному аналізі; або гіпотетично (голонасінні, покритонасінні).
Спорофіт і гаметофіт в онтогенезі не завжди є подібними і не завжди – відособленими. Тому вони становлять дві фази в онтогенезі одного покоління.
Вперше термін “чергування поколінь” в ботанічну термінологію ввів Гофмейстер.
3.2. Зміна ядерних фаз і чергування поколінь у рослин
Для кожного виду рослин, якому властиве статеве розмноження, в ядрах клітин відповідних поколінь є обов’язковий певний диплоїдний чи вдвоє менший гаплоїдний набір хромосом. Перехід від гаплоїдного до диплоїдного стану здійснюється шляхом запліднення, і, навпаки, перехід від диплоїдного до гаплоїдного забезпечується редукційним поділом ядра (мейозом). Мейоз відбувається у різних груп рослин і в різний період життя, тобто редукція може бути зиготичною, споритичною або гаметичною. Цим відрізняються більшість тварин і людина, у яких редукція хромосом відбувається обов’язково тільки перед утворенням гамет (гаметична редукція).
Для рослин відомо кілька варіантів зміни ядерних фаз, а також тривалість їх. Рослину з гаплоїдною кількістю хромосом називають гаплобіонтом, оскільки всі клітини такої рослини у вегетативному стані мають гаплоїдний набір хромосом, а рослину з диплоїдним набором – диплобіонтом. У багатьох водоростей в життєвому циклі домінуючою є гаплоїдна фаза. Такими, наприклад, є більшість зелених водоростей. Диплоїдною в них є лише зигота. Перший поділ її під час проростання, після періоду спокою, – редукційний. Отже, нова рослина, що розвивається після цього, буде гаплоїдною. Детальніше подібний цикл розвитку можна простежити на прикладі одноклітинної зеленої водорості хламідомонади (Chlamydomonas). Це одноклітинна дводжгутикова водорість кулястої, овальної або яйцеподібної, іноді несиметричної форми, з м’якою пектиновогеміцелюлозною оболонкою. Хроматофор чашоподібний з піреноїдом. Клітина має дві скоротливі вакуолі, стигму і одне гаплоїдне ядро. Розмножується вона дводжгутиковими зооспорами. При цьому клітина втрачає джгутики, а її вміст ділиться мітозом і дає 2-4 або 8 зооспор. Зооспори звільняються з оболонки материнської клітини через отвір під час її ослизнення, даючи нові дочірні, також гаплоїдні, особини.
Статеве розмноження частіше зустрічається у вигляді ізогамії, рідше – гетерогамії і оогамії.
Статеве розмноження настає за несприятливих умов. За таких умов у клітинах водорості відбувається мітотичний багатократний поділ ядра, внаслідок чого виникає багато ізогамет (32-64). Після виходу в воду ізогамети попарно копулюють. Утворена зигота, яка набула диплоїдного стану, вкривається твердою оболонкою. У зиготі накопичується достатньо запасних продуктів і червоний пігмент – гематохром. Після деякого періоду спокою (під час зими) зигота проростає, ділячись мейотично, утворюючи чотири гаплоїдні клітини, які є типовими мейоспорами. З утворенням джгутиків вони, розірвавши оболонку, випливають у воду і там розвиваються в гаплоїдні особини. Отже, в процесі відтворення, після копуляції двох гамет, сформована зигота дає чотири гаплоїдні особини, що й забезпечує більш ефективне розмноження виду.
Звертає на себе увагу і той факт, що як при нестатевому, так і статевому розмноженні дочірні особини гаплоїдні, а диплоїдною стає лише зигота.
Зовсім протилежним у ряду рослин є повне домінування в життєвому циклі диплоїдної фази (диплобіонта). Прикладом можуть бути деякі зелені, діатомові, червоні та бурі водорості. Так, у прісноводної зеленої водорості кладофори грудкуватої (Cladophora glomerata) кожна особина є диплобіонтом. Розгалужені нитки її тіла складаються з диплоїдних клітин. З настанням статевого розмноження деякі клітини ниток починають функціонувати як гаметангії. У кожному майбутньому гаметангії відбувається мейотичний поділ ядер з утворенням дводжгутикових ізогамет. Копуляція гамет відбувається у воді з наступним утворенням зиготи. Зигота проростає без. періоду спокою в нову диплоїдну особину. Для кладофори характерне також і нестатеве розмноження з участю частіше 4-джгутикових зооспор, що формуються після мітотичного поділу ядер. Після проростання вони дають диплоїдні молоді водорості. Отже, в кладофори спостерігається домінування диплоїдної фази, гаплоїдними є лише гамети.
Зміна ядерних фаз у бурої водорості фукуса (Fucus) дещо подібна до таких процесів, які є типовими для вищих тварин. Ця водорість – диплобіонт, досить складної зовнішньої і внутрішньої будови. У рослини, яка досягла статевої зрілості, у окремих спеціальних заглибинах утворюються, жіночі статеві органи – оогонії, в інших чоловічі – антеридії, в яких формуванню гамет передує мейотичний поділ. Утворені яйцеклітини і сперматозоїди гаплоїдні. Після виходу їх із статевих органів у воду відбувається запліднення яйцеклітин сперматозоїдами. Кожна утворена зигота вкривається оболонкою і відразу розвивається в нову диплоїдну особину фукуса (мал.22). Отже, фукус все своє життя, як і організми вищих тварин, перебуває в диплофазі, гаплофаза обмежена лише часом існування гамет. Немає також і спороношення.
Отже, у процесі життя рослин спостерігається явище переходу від гаплоїдного стану до диплоїдного і навпаки, воно зумовлене зміною ядерних фаз. Слід зазначити, що зміна ядерних фаз не залежить від рівня морфологічної організації особин чи особливостей їхньої життєдіяльності, вона властива всім нижчим еукаріотам і всім вищим рослинам.
3.3. Чергування поколінь
У багатьох водоростей і всіх вищих рослин чергування гаплофази і диплофази у життєвому циклі пов’язане з більш складним процесом чергування поколінь – гаплобіонтів і диплобіонтів. Кожне покоління відрізняється від свого попереднього насамперед кількістю хромосом, а часто і розмірами, а також зовнішнім виглядом.
Рослина в диплофазі розмножується нестатево – спорами, ут-ворення яких супроводжується редукційним поділом. Із спор роз-вивається гаплоїдна рослина – гаметофіт – з наступним утворенням гамет. Гамети після злиття утворюють зиготу. Остання розвивається в нову диплоїдну рослину. При цьому спостерігається не лише зміна гаплоїдної і диплоїдної ядерних фаз, а й відповідне їм чергування поколінь – статевого (гаплоїдного) і нестатевого (диплоїдного). Нерідко кожне з поколінь веде самостійне життя і розмножується статевим або нестатевим способом. Статеве покоління називають гаметофітом.
За своєю зовнішньою формою гаметофіти і спорофіти в тих самих водоростей подібні, в інших досить відрізняються між собою, крім одноклітинних, у яких життєві форми однакові. До першої групи можна віднести морську зелену водорість ульву (Ulva). У неї статеве і нестатеве покоління однаково добре розвинуті і зовні (морфологічно) відрізнити не можна. Слань має вигляд двошарової багатоклітинної по краях складчастої пластинки, звуженої у коротку ніжку, якою прикріплюється до субстрату. У циклі розвитку ульви розрізняють нестатеву рослину (спорофіт), в якій розвиваються тільки зооспори, і статеву (гаметофіт), яка морфологічно схожа з першою, але в ній розвиваються тільки зооспори. Зигота, утворена після копуляції гамет, без періоду спокою і без редукційного поділу, проростає в диплоїдну рослину – спорофіт. Утворенню зооспор передує редукційний поділ. Зооспори після дозрівання і звільнення із зооспорангіїв проростають, даючи початок гаплоїдному статевому поколінню – гаметофіту, який розвиває гаметангії і гамети. Такий тип чергування або зміни поколінь у циклі розвитку, коли морфологічно подібні як спорофіт, так і гаметофіт, але відрізняються фізіологічно і цитологічно, називають ізоморфним.
Ізоморфне чергування поколінь вважається стародавнім і пер-винним за походженням, яке яскраво підтверджує, що обидві життєві фази (спорофіт і гаметофіт) того самого виду почали існувати як рівнозначні і незалежні в тих самих умовах.
У процесі історичного розвитку в деяких водоростей спостері-галася морфологічна редукція одного з поколінь: в одних редуку-валось диплоїдне покоління, в інших – гаплоїдне. Редукція одного з поколінь, як підтверджують дослідження І.Перро (1972), тісно пов’язана із зміною зовнішніх умов. Прикладом першої еволюції гілки, коли переважаючим (домінуючим) і відповідно більш морфологічно розвинутим є гаплобіонт, може бути улотрикс (Ulotrix zonata, мал. 23), який зустрічається в різних прісноводних водоймах і гірських річках. Це нерозгалужена однорядна нитчаста водорість, прикріплена базальною безбарвною клітиною до субстрату. Всі клітини нитки, крім базальної, здатні до поділу і розмноження, в кожній з них є один пояскоподібний хроматофор, кілька піреноїдів, одне ядро і центральна вакуоля, заповнена клітинним соком. Це гаплобіонт. Він розмножується нестатево – мікро- і макрозооспорами, які несуть 4, рідше 2, джгутики, що утворюються мітозом. Проростаючи, вони дають такі ж гаплоїдні особини. Статеве розмноження – ізогамія, обов’язковим є гетероталізм (копулюють гамети різних особин). Утворена зигота дає одноклітинне гаплоїдне покоління – спорофіт. Ця диплоїдна мікроскопічна водорість, що має вигляд округлої клітини на короткій ніжці (дослідження Ю.Є.Петрова, П.Корнман), після дозрі-вання і мейотичного поділу утворює зооспори. Після деякого часу руху зооспори втрачають джгутики, осідають і, проростаюч, утворюють гаплоїдні нитчасті гаметофіти. Такий цикл розвитку серед нижчих рослин досить поширений. Він характерний для багатьох зелених (улотриксові, едогонієві), бурих і червоних водоростей.
Напрям, коли в життєвому циклі більш редуковане гаплоїдне покоління, в нижчих рослин зустрічається значно рідше. Як приклад цього напряму доцільно розглянути цикл розвитку відомої бурої водорості – ламінарії.
Ламінарія порівняно з іншими водоростями досягла високого рівня морфологічної організації. Її спорофіт є багаторічною, великою (до 6 м) рослиною, розрізняють розчленування талому на ризоїдальну, стеблову (каулоїд) і листкоподібну (філоїд) частини. Форма талому має вигляд суцільної або розсіченої пластинки. Наростає він у результаті діяльності меристеми, яка розташована на межі стебло- і листкоподібної частини (інтеркалярно). Внут-рішня будова тканинна.
На філоїдах спорофіта з периферійних клітин утворюються соруси одноклітинних зооспорангіїв, в яких після редукційного поділу розвиваються численні дводжгутикові зооспори. Після виходу у воду вони осідають на дно і проростають, утворюючи гаплоїдні одностатеві гаметофіти (заростки), що мають вигляд коротких розгалужених ниток. Після деякого часу дозрівання чоловічий гаметофіт з окремих клітин бічних гілок утворює антеридії з чис-ленними сперматозоїдами. В цей час на жіночому гаметофіті, на кінчиках розгалужень ниток, утворюються оогонії з яйцеклітинами Запліднення відбувається у воді. З утвореної зиготи без періоду спокою розвивається диплоїдна особина – спорофіт. Отже, спорофіт ламінарії є домінуючим поколінням. Порівняно з гаметофітом він значно більший (до 6 м) і досить складної будови. Гаметофіт, як чоловічій, так і жіночій, – мікроскопічно малий, нит-коподібний. Він живе протягом 2,5 року. За цей час відбувається формування його, утворення статевих органів, гамет і навіть вегетативне розмноження.
Розглянуті вище напрямки розвитку спорофіта і гаметофіта у водоростей продовжуються і в наземних (вищих) рослин. Так, у мохів домінує гаметофіт. Це найпримітивніші сучасні наземні рослини.
Цикл розвитку мохоподібних розглянемо на прикладі зеленого моху зозулиного льону звичайного (Рolytrіchum commune).
Його гаметофіт дводомний, має листкостеблову будову, прикріплюється до субстрату за допомогою багатоклітинних розгалужених ризоїдів (мал. 24). На верхівках чоловічих рослин в оточенні червоно-бурнх листкових розеток утворюються антеридії (чоловічі гаметангії), що мають вигляд округлих або видовжених мішечків. В них із сперматогенних клітин формуються рухомі дводжгутикові сперматозоїди.
На верхівках жіночих рослин в оточенні зелених листочків утворюються архегонії (жіночі гаметангії). Архегоній, на відміну від оогонія, має колбоподібну форму з розширеним черевцем, яке містить велику яйцеклітину, а над нею черевну канальцеву клітину і звужену довгу шийку з одним рядом канальцевих клітин. З дозріванням архегонія шийкові і черевні канальцеві клітини ослизнюються і утворюється вузький канал, виповнений рідиною, за допомогою якої сперматозоїд може проникнути до яйцеклітини.
Органи статевого розмноження моху (антеридії і архегонії) багатоклітинні і захищені від несприятливих умов багатоклітинним зовнішнім покривним шаром. Оскільки у водному середовищі гаметам не погрожувало висихання, з переходом рослин до наземних умов у гаметангіях виробилось відповідне пристосування, що й виражене в їх багатоклітинності. Гаметангії, формуючись найчастіше групами, швидше накопичують і краще зберігають вологу. Однак океан був первородним середовищем для рослин протягом довгого часу і втратити зв’язок з водним середовищем, особливо в процесі розмноження, вони відразу не змогли. Для запліднення у мохоподібних обов’язковою умовою є вода. Під час дощу або роси антеридії розкриваються і випускають численні сперматозоїди. Рухаючись у воді, вони притягуються речовинами, які архегоній виділяє разом із слизом, проникають через каналець шинки до яйцеклітини. Злиття гамет і подальший розвиток зиготи відбувається всередині черевця архегонія. Незважаючи на те, що в більшості мохів розвивається кілька архегоніїв, але запліднюється лише один.
На верхівці жіночого гаметофіта в заплідненому архегонії із зиготи розвивається спорогоній (від гр. spora – посів, honos – народження) – спорофіт. Він простої будови. Складається з одного спорангія – коробочки, розташованої на ніжці, що в нижній частині переходить в присоску, за допомогою якої вростає в тканину гаметофіта. Коробочку прикриває дуже змінений архегоній – ковпачок. Клітини спорогонія диплоїдні. В його спорангії з клітин археспорія (від гр. arhe – початок, spora – посів) після редукційного поділу формуються спори, які, вкрившись твердою оболонкою, висипаються. В сприятливих умовах із спори розвивається розгалужена нитчаста протонема (від гр. protos – перший, nema – нитка), яка в більшості мохів зовні схожа на нитчасту зелену водорість. Це зумовило не підтверджену думку про походження мохоподібних від зелених водоростей. На протонемі закладається кілька бруньок, з яких розвиваються нові гаплоїдні рослини – гаметофіти.
Таким чином, поряд з виникненням статевого ризмноження, гаметофіт виконує ще й функції фотосинтезу, водозабезпечення, мінерального живлення, а також вегетативного розмноження. Спорофіт же, розвиваючись на гаметофіті, фактично обмежений тільки спороутворенням.
Повний життєвий цикл від зиготи до зиготи складається із гаметофіта, розвиток якого починається від спори і закінчується утворенням гамет, і спорофіта, який бере свій початок із зиготи і закінчується утворенням спор статевого розмноження.
3.4. Чергування поколінь у рівноспорових папоротей
В усіх інших вищих рослин домінуючим, відповідно більш розвинутим, є спорофіт (диплобіонт). Щоб вивчити поставлене питання про чергування поколінь у рівноспорових папоротей, доцільно більш детально розглянути на прикладі звичайної папороті чоловічої, або щитника чоловічого (Dryoppteris filix mas, мал. 25). Нестатевим поколінням (спорофітом) у щитника є сама рослина, з кореневищем (підземним пагоном), коренем і листками – вайями. Ядра клітин мають диплоїдний набір хромосом. З нижнього боку листків на потовщеннях – плацентах (від лат placenta – перепічка, орган зв’язку зародка з материнським організмом) закладаються спорангії. Вони округлої форми, але дещо сплющені з боків, сидять на коротких ніжках. У більшості папоротей спорангії розвиваються на звичайних листках, а в деяких – на видозмінених, що називають спорофітами. У багатьох папоротей спорангії розміщуються на плацентах групами – сорусами (від гр. soros – купа, група спор). Кожен сорус зверху покритий ідузіумом (від лат. indusium – покривальце), (покривальцем), що виростає з плаценти.
Кожен спорангій розвивається з меристематичного горбочка. Причому зовнішні його клітини утворюють оболонку спорангія, а внутрішні – археспоріальну тканину. Зовнішні клітини археспорангія дають початок вистилаючому шару, або тапетуту (від гр. tapetum – покривало, килим), а внутрішні після деяких мітотичних поділів – спорогенній тканині. Клітини спорогенної тканини багато разів діляться мітотичнo (останній поділ їх буде мейотичний) з утворенням тетради гаплоїдних клітин, які після утворення оболонок роз’єднуються. Це й є спори статевого розмноження. Дозрілі спори мають дві оболонки. Зовнішня (екзина) товста і бугриста, темно-бурого забарвлення, а внутрішня (інтина) тонка, еластична.
Розкриття спорангія і звільнення дозрілих спор відбувається за допомогою фіброзного шару, в якого клітини мають нерівномірно потовщені оболонки При зневодненні фіброзне кільце випростовується і розриває протилежну стінку спорангія з тонкостінних клітин і спорангій відкривається. Виштовхнуті із спорангія спори висипаються. Розповсюджуються вони вітром або водою і, потрапляючи в сприятливі умови, проростають, утворюючи заростки. Вони мають вигляд багатоклітинних маленьких (2-5 мм діаметром кожна) зелених серцеподібної форми пластинок. До грунту пластинки прикріплюються ризоїдами. Тіло кожного заростка (гаметофіта) складається з гаплоїдних клітин і є гаплобюнтом. Гаплобіонтом морфологічно зовсім не схожий з диплоїдною рослиною – диплобіонтом (спорофітом).
Через деякий час на нижній частині заростка виникають гаметангіі – спочатку антеридії, потім архегонії. В антеридіях і архегоніях з редукцією кількості хромосом формуються гамети-сперматозоїди і яйцеклітини.
У більшості папоротей гаметофіти є наземними, асимілюючими двостатевими формами.
Антеридії папороті, що розглядається, мікроскопічні, за формою мішкоподібні, з одношаровою стінкою. Всередині кожного антеридія розвиваються сперматогенні клітини, з яких виникають сперматозоїди, які мають джгутики.
Архегоніі, як і в інших спорових рослин, колбоподібної будови і теж з одношаровою стінкою. Розширена частина архегонія – черевце, заглиблене в тканину заростка, а звужена його частина – шийка, що більш-менш висовується назовні. У черевці знаходиться яйцеклітина з великим ядром і черевна канальцева клітина, шийка вміщує кілька шийкових канальцевих клітин. З дозріванням архегонія шийка розкривається, канальцеві клітини ослизнюються і утворена слизиста речовина виходить у воду і хімічно діє на сперматозоїди, що вийшли з антеридіїв. Рух сперматозоїдів до архегоніїв і проникнення їх до черевця, де міститься яйцеклітина, підкоряється законам хемотаксису (тут діє одна з форм таксисів). З прониклих сперматозоїдів лише один зливається з яйцеклітиною, запліднюючи її. Решта сперматозоїдів гинуть. Запліднена яйцеклітина вкривається оболонкою і ділиться мітотично. Процес поділу і розвиток зиготи відбувається досить інтенсивно, і незабаром виникають усі частини зародка, властиві папороті: корінь, первинний листок, стебло і ніжка, якою зародок прикріплюється до заростка. Перші листки найчастіше не схожі з листками дорослої папороті, але згодом ця відмінність зникає. Молодий зародок спочатку розвивається за рахунок запасів поживних речовин, що є в заростку (гаметофіті), але через деякий час він руйнується і відмирає. В цей час молода рослина повністю переходить до самостійного живлення.
У папороті закономірно змінюються не лише ядерні фази, межами яких є утворення мейоспор (перехід від диплофази до гаплофази) і статевий процес (перехід від гаплофази до диплофази), а й два різних за виглядом і функціями самостійних покоління рослин: нестатеве (диплоідний спорофіт) і статеве (гаплоїдний гаметофіт). Відтворення собі подібного відбувається лише через одне покоління.
У папоротей спорофітом є багаторічна рослина з добре розвиненими вегетативними органами, яка відповідно пристосована до життя в наземних умовах. Гаметофіт – невелика, майже мікроскопічна, таломна, короткочасна рослина, пристосована до виконання статевого розмноження. Тобто обидва покоління морфологічно, цитологічно і за своїми функціями різні. Таку зміну поколінь називають гетероморфною. У папороті кожна зигота здатна розвиватися в спорофітну рослину, яка утворює мільйони мейоспор. Тому біологічне значення нестатевого покоління полягає у підвищенні здатності виду до розмноження і розселення.
3.5. Апоміксис, його роль у житті квіткових рослин
Крім звичайного подвійного запліднення у квіткових рослин часто спостерігається відхилення від нормального статевого процесу, коли зародок і насіння розвиваються без запліднення. Такий пооцес називають апоміксисом (від гр. аро – без, mixis – змішування).
Існують такі категорії апоміксису: партеногенез (від гр. раrthenos – незаймана, genesis – походження), апогамія (від гр. аро – без, gamos – брак), апоспорія (від гр. аро – без, spora – сіяння). Наведена класифікація базується на основі того, з яких компонентів насінного зачатка розвивається зародок. Якщо він утворюється з незаплідненої яйцеклітини, таке явище називають партеногенезом. Розвиток партеногенезу відбувається двома шляхами. В одних випадках зародок формується з гаплоїдної яйцеклітини (у дурману, тютюну та ін.), в інших – з диплоїдної. Причому диплоїдний набір хромосом зберігається, бо в процесі розвитку зародкового мішка не відбувся мейоз і яйцеклітина залишилася диплоїдною. Так розвивається насіння у манжеток, нечуйвітру, кульбаби.
Явище апогамії характеризується тим, що зародок насінини формується не за рахунок яйцеклітини. У цьому процесі участь беруть синергіди, антиподи (в редукованому або нередукованому стані), а також клітини ендосперму. Апогамія спостерігається у подорожника ланцетолистого, цибулі пахучої та інших рослин.
Третій вид апоміксису апоспорія проявляється як особлива його форма, коли зародок розвивається поза гаметофітною фазою за рахунок диплоїдних клітин нуцелуса, халази або покривів насінного зачатка. Апоспорія спостерігається у цитрусових, розоцвітих та ін. Виділяють два варіанти апоспорії.
1. Нормальний зародковий мішок дегенерує. Зародок формується з диплоїдних клітин нуцелуса або халази. Утворені клітини зародка або безпосередньо вростають всередину відмираючого зародкового мішка, або з нуцелярних чи халазних клітин розвивається новий зародковий мішок з зародком, і весь цей утвір продовжує розвиватися у дегенерованому мішку. Таким чином, у даному випадку зародок формується з деякою участю нормального зародкового мішка.
2. Зародок розвивається за рахунок клітин нуцелуса, а інколи й покривів без будь-якого зв’язку із зародковим мішком і носить характер додаткової, або адвентивної, ембріонії. Такий вид апоміксису часто проявляється у мандарина, лимона, апельсина і зумовлює багатозачатковість (поліембріонію), внаслідок чого утворюється до 20 зародків, але нормально розвивається 1-4, решта відмирають.
Характерною особливістю такої ембріонії є те, що тут спостерігається своєрідний перехід між статевим і вегетативним розмноженням, зародок формується за рахунок диплоїдних клітин спорофіта, а гаметофітний стан взагалі випадає з цик- лу розвитку.
Розглядаючи весь процес розвитку апоміксису, можна характеризувати його як процес вегетативного розмноження, бо він відбувається без запліднення. У багатьох рослин апоміктичний шлях розвитку насіння став звичайним, і рослини добре пристосовувалися до розмноження, заселяючи інколи великі території. Налічують майже 50 родин квіткових рослин, у яких спостерігається апоміксис, але найбільше поширення він має серед жовтецевих, орхідних, лілійних, злаків та інших родин рослин.
22. Хрящові риби (Chondrichthyes) — клас хребетних. Відомі із середнього девону. Довжина від 6 см до 20 м. Кістяк хрящовий, часто затверділий (обвапнований). Тіла хребців і ребра є або відсутні. Шкірних кісток немає. Луска, якщо є, плакоїдна. Лопасті плавців підтримуються еластиновими нитками. 5—7 зябрових щілин, що відкриваються назовні, в деяких 4 щілини, прикриті шкірною складкою. Плавального міхура немає. Кишечник зі спіральним клапаном; в серці є артеріальний конус. Анальний і сечостатевий отвори біля основи черевних плавців. Запліднення внутрішнє. Статеві органи в самців (птеригоподії) парні, розвиваються з черевних плавців. Живородні (рідко), яйцеживородні або відкладають великі яйця. Осмотичний тиск внутрішнього середовища в хрящових риб забезпечується головним чином за рахунок сечовини, розчиненої в крові. Порожнинні рідини гіпертонічні стосовно навколишнього середовища. При перенесенні хрящових риб в прісну воду осмотичний тиск крові й інших порожнинних рідин падає і вони швидко гинуть. Прісноводні хрящові риби для збереження високого осмотичного тиску виділяють велику кількість сечі. 2 сучасних підкласи — пластинчатозяброві і суцільноголові; близько 140 сучасних родів і близько 630 видів. Живуть в морі (кілька видів — у прісних водах).
Біологічна класифікація
Домен: Ядерні (Eukaryota)
Царство: Тварини (Metazoa)
Підцарство: Справжні багатоклітинні (Eumetazoa)
Тип: Хордові (Chordata)
Підтип: Черепні (Craniata)
Інфратип: Щелепні (Gnathostomata)
Клас: Хрящові риби (Chondrichthyes)
Хрящові риби мають ряд прогресивних ознак порівняно з круглоротими та безчерепними:
парні грудні та черевні плавці
хрящовий внутрішній скелет
щелепи
внутрішнє запліднення
селезінка
підшлункова залоза
Форма тіла та покриви
Форма тіла в акул торпедоподібна, у скатів — сплющена в спинно-черевному напрямі. Є плавці: парні черевні та грудні, непарні спинний і хвостовий. Для хрящових риб характерна плакоїдна луска, що складається з дентинової пластинки, розташованої в дермі, та вкритого тонким шаром емалі зубця, який пронизує епідерміс. Шкіра хрящових риб виділяє слиз, який запобігає проникненню крізь покриви бактерій і зменшує тертя під час руху.
Скелет
Скелет утворений хрящовою тканиною, часто насиченою солями кальцію, що надає їй міцності. Скелет підрозділяється на хребет, череп, скелет плавців (кінцівок) та їхніх поясів. Хребет складається з хребців, має два відділи: тулубний і хвостовий. Хорда зберігається протягом усього життя у вигляді тонкого тяжа в центрі тіл хребців і заповнює простір між ними. Тіла хребців тулубного відділу утворюють верхні дуги, через які проходить спинний мозок. У хвостовому відділі хребці мають ще нижні дуги, через які проходити хвостова артерія та вена. Хвіст — найрухливіша частина тіла риб, тому його судини потребують захисту від здавлювання.
Череп складається з двох відділів — мозкового та вісцерального. Мозковий череп повністю прикриває головний мозок. Вісцеральний череп утворений зябровими дугами — опорою для дихального апарату, і щелепами — зябровими дугами, видозміненими у зв'язку з активним способом життя.
Пояс передніх кінцівок (плечовий) представлений хрящовою дугою, пояс задніх кінцівок (тазовий) — непарною хрящовою пластинкою. Вільні плавці утворюються з рядів хрящів.
Мускулатура
Мускулатура хрящових риб складається з окремих поперечносмугастих м'язових сегментів.
Нервова система й органи чуттів
Головний мозок хрящових риб має ті самі п'ять відділів, що й круглороті, проте кількість нервових клітин у кожному з них істотно зростає. Великого розвитку досягають передній мозок і мозочок. З органів чуттів провідним є орган нюху. Він складається з парних нюхових мішків, що відкриваються назовні ніздрями. У мішки надходить вода, і розчинені в ній молекули речовин подразнюють чутливі клітини. Очі в акул розвинені добре, дозволяють розрізняти контури предметів на відстані 10—15 м. Орган слуху представлений внутрішнім вухом, орган рівноваги — трьома напівкруглими каналами. Бічна лінія складається зі скупчення чутливих клітин на тулубі та голові. Кожна така клітина має виріст, який сприймає рух води.
Травна система
Більшість акул — хижаки. Органом захоплення й утримання здобичі є щелепи. Вони наділені кількома рядами гострих зубів різноманітної форми. Якщо один зуб зламається, він замінюється іншим. За своєю природою зуби є збільшеними лусками. З ротової порожнини їжа потрапляє до глотки, яка має зяброві щілини. З глотки харчові частинки рухаються в короткий стравохід і далі в об'ємний шлунок. Тут їжа поволі (до 5 діб) перетравлюється. Далі йде кишечник (тонка, товста та пряма кишки), де відбуваються завершальні етапи травлення і процес всмоктування. Товста кишка має внутрішній виріст — спіральний клапан, що значно збільшує всмоктувальну поверхню. У просвіт кишки відкриваються протоки печінки та підшлункової залози. Неперетравлені залишки через пряму кишку потрапляють у клоаку та виводяться назовні.
Дихальна система
Органами дихання є зябра. У більшості акул і скатів глотка пронизана п'ятьма парами зябрових щілин. Кожна щілина утворена зябровою перегородкою, прикріпленою до відповідної хрящової зябрової дуги. Зябра являють собою зяброві пелюстки, розташовані по обидва боки від зябрової перегородки. Вода постійно омиває зяброві пелюстки, густо обплетені капілярами.
Кровоносна система
Кровоносна система замкнена. Серце велике, складається з одного передсердя та шлуночка з товстими м'язовими стінками. До передсердя примикає венозний синус (у круглоротих відсутній), куди збирається венозна кров із усього організму. Як і в круглоротих, через серце хрящових риб проходить тільки венозна кров. Скороченнями шлуночка вона просувається до капілярів зябер, де збагачується киснем. Далі кров збирається в спинну аорту й розноситься нею по всьому тілу. У хвостовому відділі вона входить у канал, утворений нижніми дугами хребців, і називається хвостовою артерією.
Видільна система
Система виділення хрящових представлена парою тулубних нирок. Кожна нирка містить безліч нефронів, що складаються з капсули та канальців. У нефроні відбувається фільтрація та реабсорбція потрібних організму речовин. Канальці зливаються з утворенням сечоводу, який відкривається в клоаку.
Статева система та розвиток
Статеві залози парні. У самців сім'яники мають власні вивідні протоки — сім'япроводи, які відкриваються в клоаку. Черевні плавці у самців акул виконують функцію купулятивного органа. У самок яйцеклітини з яєчника потрапляють в яйцепроводи, де й відбувається внутрішнє запліднення. Яйця відкладаються на різні субстрати; у більшості видів вони добре захищені: проходячи яйцепроводами, запліднена яйцеклітина одягається шкаралупою. Це дає можливість хрящовим відкладати невелику кількість яєць (1—10), збагачених жовтком.
Розвиток прямий. Іноді спостерігається яйцеживородіння (у катрана), коли ембріон розвивається за рахунок запасів яєчного жовтка, але в яйцепроводах матері. Деякі види (скат-хвостокіл, акула-молот) є живородящі. У них утворюються вирости яйцепроводу, через які зародок одержує поживні речовини з організму матері.
Екологія та різноманітність
Описано близько 350 видів акул, розміром від 15 см (плоскохвоста акула) до 15—20 м (китова акула). Акули значно поширені в морях і океанах, зустрічаються навіть у прісній воді. Більшість — хижаки, але китова та гігантська акули живляться тільки планктоном. Акули не мають плавального міхура і повинні весь час плисти, щоб не потонути. Крім того, тільки за досить великої швидкості їхні зябра омиваються необхідною для дихання кількістю води. У сучасних дослідженнях називається 50 видів акул, потенційно небезпечних для людини.
23.Ядро. У клітинній біології, ядро (лат. nucleus) – клітинна органела, знайдена у більшості клітин еукаріотів і містить ядерні гени, які складають більшу частину генетичного матеріалу. Ядро має дві первинні функції: керування хімічними реакціями в межах цитоплазми і збереження інформації, потрібної для поділу клітини.
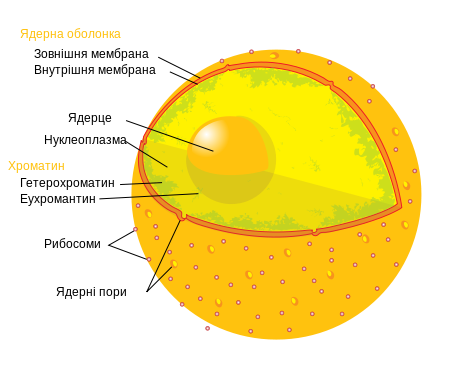
Окрім клітинного геному, ядро містить певні білки, які регулюють зчитування генетичної інформації. Зчитування гена на ядерному рівні залучає складні процеси транскрипції, обробки первинної мРНК і експорт зрілої мРНК до цитоплазми.
Ядро звичайно має розмір 8-25 мікрометрів в діаметрі. Воно оточено подвійною мембраною, яка називається ядерною оболонкою. Крізь внутрішню і зовнішню мембрани на деяких інтервалах проходять ядерні пори. Ядерна оболонка регулює і полегшує транспорт між ядром і цитоплазмою, відокремлюючи хімічні реакції, що відбуваються в цитоплазмі, від реакцій, що трапляються в межах ядра. Зовнішня мембрана безперервна з грубим ендоплазматичним ретикулумом (англ. RER) і може мати зв’язані рибосоми. Простір між двома мембранами (який називається «перинуклеарним простором») безперервний з люменом RER. Ядерна сторона ядерної оболонки оточена мережею філаментів, яка називаються ядерною ламіною.
Внутрішня частина ядра містить одне або декілька ядерець, оточених матрицею, яка називається нуклеоплазмою. Нуклеоплазма (каріолімфа, ядерний сік, каріоплазма) – гелеподібна рідина (подібна у цьому відношенні до цитоплазми), в якій розчинені багато речовин. Ці речовини включають нуклеотид-тріфосфати, сигнальні молекули, ДНК, РНК та білки (ензими та філаменти).
Ядро та ендомембранна система
Генетичний матеріал присутній в ядрі у вигляді хроматину, або комплексу білка і ДНК. ДНК присутня як цілий ряд дискретних молекул, відомих як хромосоми. Є два види хроматину: еухроматин і гетерохроматин. Еухроматин - менш компактна форма ДНК, області ДНК які знаходяться у формі еухроматину містять гени, які часто зчитуються клітиною.
У гетерохроматині ДНК більш компактно упакована. Області ДНК які знаходяться у формі гетерохроматину містять гени, які не зчитуються клітиною на даній стадії розвитку (цей вид гетерохроматину відомий як факультативний гетерохроматин) або є областями, які складають теломери і центромери хромосом (цей вид гетерохроматину відомий як конструктивний гетерохроматин). У багатоклітинних організмах, клітини надзвичайно спеціалізовані, щоб виконувати специфічні функції, тому різні набори генів потрібні і зчитуються. Тому, області ДНК, які знаходяться у формі гетерохроматину, залежать від типу клітини.
Ядерце - щільна структура в ядрі, де збираються елементи рибосом. У ядрі може бути одне або декілька ядерець.
Історія відкриття
В 1831 році англійський природознавець Роберт Браун вивчав різні види рослин, зразки яких він зібрав під час подорожі до Австралії. Браун був дуже уважним до деталей, а клітини рослин особливо цікавили його. Розглядаючи їх під мікроскопом, він побачив дещо цікаве: кожна клітина містила круглий і непрозорий елемент. Він назвав його ядром.
Дізнавшись про спостереження Брауна, німецький фізіолог Теодор Шванн почав шукати подібні елементи в клітинах пуголовків і знайшов. Кожна клітина містила ядро. Це був революційний прорив - свідчення того, що всі види життя пов'язані між собою. В одній із книг Шванн описав різні типи клітин, взяті від різноманітних організмів і визначив їх за фактом наявності ядра.
Усвідомлення того, що є елемент спільний для всіх організмів, не тільки для рослин, а й для тварин, поєднало рослинне і тваринне царство у щось спільне, щось, що мало однакові риси...
24.Еволюційна теорія — наукова теорія, що пояснює поступальний розвиток життя на Землі від простих форм (клітини, бактерії) до складних. У біології, еволюція — це зміна в популяції успадкованих характеристик або ознак, які передаються з роду в рід. Ці характеристики кодуються в генах, копіюються і передаються до наступних поколінь при розмноженні. Випадкові зміни в генах можуть спричиняти нові або змінені ознаки, призводячи до відмінностей між організмами. Процес еволюції відбувається, коли різні ознаки стають загальнішими або переважаючими в популяції. Це може відбуватися безладно через генетичний дрейф, або шляхом природного добору через боротьбу за існування та виживання найпридатніших.
При природному доборі, організми з ознаками, які допомагають їм вижити і відтворитися, як правило, мають більше нащадків. Таким чином, вони передадуть більше вигідних спадкових ознак наступним поколінням. Це призводить до того, що вигідні ознаки стають загальнішими в кожному поколінні, а невигідні зменшуються. Через деякий час, цей процес «мікроеволюції» може приводити до різноманітних адаптацій в залежності від замін в навколишньому середовищі. Коли відмінності всередині та між популяціями накопичуються, це приводить до утворення нових видів і вищих таксонів (макроеволюції). Всі відомі види так чи інакше походять від єдиного предка через процес поступової розбіжності.
Теорія еволюції шляхом природного добору була вперше запропонована в книзі Чарльза Дарвіна «Походження видів» 1859 р. У 1930-х, теорія природного добору Дарвіна разом з теорією наслідування Менделя склали сучасну синтетичну теорію еволюції. З її величезними властивостями пояснення та передбачення, ця теорія стала центральним організовуючим принципом сучасної біології та соціології, забезпечуючи загальне пояснення різноманітності життя на Землі й появи ноосфери.
Ч. Дарвін
Еволюційна теорія англійського ученого Ч. Дарвіна (1809—1882) має основні положення:
1. Умови зовнішнього середовища викликають зміну організмів, але ці зміни (модифікації) не успадковуються і не відіграють важливої ролі в еволюції.
2. Випадкові, ненаправлені зміни в окремих особин (мутації) можуть передаватися по спадку, якщо вони відбуваються в статевих клітинах. Саме такі зміни важливі для еволюції.
3. Усі успадковані зміни відносні, тобто в одних умовах вони можуть бути корисними, в інших — шкідливими або нейтральними. Якщо в даних умовах вони виявляються корисними, то організми з такими змінами виживають і дають потомство.
Головні рушійні сили еволюції: боротьба за існування; природний добір на основі спадкової мінливості.
Основні чинники еволюції: спадкова мінливість; природний добір; ізоляція; популяційні хвилі; дрейф генів.
Результати еволюції: відносна пристосованість видів; ускладнення організації; видоутворення.
Боротьба за існування — це активна або пасивна конкуренція між організмами, що виникає через невідповідність між здатністю видів до необмеженого розмноження і обмеженістю природних ресурсів. Розрізняють такі види боротьби за існування: внутрішньовидова, міжвидова, боротьба з несприятливими умовами середовища. Внутрішньовидова боротьба викликається однаковою у всіх особин даного виду потребою в їжі, території та інших ресурсах. Міжвидова боротьба виникає між особинами різних видів, що використовують одні й ті ж харчові ресурси, територію. Боротьба з несприятливими умовами середовища полягає у виживанні організмів при постійній дії абіотичних чинників середовища — повеней, засух, морозів, приливів і відливів, штормів, заболочування, сніжної лавини, селів, пожеж і т. д.
Результатом боротьби за існування є виживання найбільш пристосованих і їх розмноження. Слабкі особини гинуть або не породжують потомства, їх гени елімінуються. У ході боротьби за існування відбувається природний добір — вибіркове знищення одних особин і вибіркове виживання інших.
Природний добір — процес, у результаті якого найбільш пристосовані організми виживають і розмножуються, а менш пристосовані — гинуть і не залишають потомства. Природний добір — це єдиний чинник еволюції, що здійснює направлену зміну генотипного складу популяцій. Виділяють три типи добору — направлений, стабілізувальний і дизруптивний (розриваючий).
Направлений, або рушійний, добір викликає однонаправлені зміни генетичного складу популяції. Він спостерігається в тих випадках, коли популяція пристосовується до нового середовища. Стабілізувальний добір не сприяє еволюційним змінам і з покоління в покоління підтримує фенотипну стабільність популяції. Якщо популяція добре пристосована до стабільного середовища, то добір зводиться до елімінації мутантних особин і особин, які відхиляються від норми.
Дизруптивний (розриваючий) добір направлений проти особин із середнім і проміжним характером ознак, тобто веде до стабілізації крайніх форм і встановлення поліморфізму в популяції. Популяція при цьому підрозділяється за даною ознакою на декілька груп.
Відносна пристосованість
Результатом дії рушійних сил еволюції є відносна пристосованість видів до умов навколишнього середовища. У поняття «пристосованість» входять забарвлення, форма тіла, поведінка, особливості фізіологічних і біохімічних процесів і т. д.
Одним з найяскравіших прикладів пристосованості є захисне забарвлення, що робить тварину схожою з навколишніми предметами. Деякі тварини можуть змінювати забарвлення тіла під колір місцевості, наприклад хамелеони і восьминоги. Різновид захисного забарвлення — розчленовуюче, як у тигра і зебри. Чергування різнокольорових вертикальних смуг розбиває контур тварини на фоні навколишнього пейзажу та інших особин стада. Деякі тварини мають не тільки захисне забарвлення, але й маскуючу форму тіла. Захисне (загрозливе) забарвлення попереджає хижаків, що її власник отруйний або неїстівний. Це різке, яскраве забарвлення, як, наприклад, у сонечка. Деякі беззахисні види мають забарвлення, схоже з таким у неспоріднених, добре захищених і маючих захисне забарвлення видів. Таке наслідування називається мімікрією. Пристосувальна поведінка має велике значення для виживання організмів у боротьбі за існування. Існують різні варіанти пристосувальної поведінки: затаювання у разі небезпеки, демонстративна і відлякуюча поведінка у разі наближення ворога, запасання корму на несприятливий сезон, різноманітні форми піклування про потомство.
Дрейф генів — випадкові, ненаправлені зміни частот алелей у популяції. Роль дрейфу генів особливо зростає, коли чисельність популяції різко скорочується (у результаті катастроф, зміни середовища, популяційних хвиль). При цьому через випадковість у популяції можуть значно зрости або знизитися частоти тих або інших алелей. Через зростання частки близькоспоріднених схрещувань деякі алелі можуть повністю зникнути з популяції, а інші, навпаки, стати переважаючими.
Популяційні хвилі (коливання чисельності особин, що становлять популяцію) — це чинник еволюції, який припускає невиборче, випадкове знищення особин, завдяки чому рідкісний генотип (алель) може стати звичайним і підхопитися природним добором.
Причини коливання чисельності популяцій:
1) періодичні (сезонні) коливання чисельності характерні для короткоживучих організмів — комах, найпростіших, однорічних рослин;
2) неперіодичні коливання чисельності залежать від складного поєднання ряду чинників — чисельності хижаків, великої кількості кормових ресурсів;
3) спалахи чисельності видів спостерігаються в нових для них районах, де відсутні їхні природні вороги (кролики в Австралії, домові горобці в Північній Америці);
4) різкі коливання чисельності пов’язані з природними катастрофами (засухою, пожежами, повенями та ін.).
У популяції, що пережила катастрофічне скорочення чисельності, частоти алелей будуть іншими, ніж у початковій популяції. При цьому деякі присутні в малих концентраціях алелі можуть зникнути з популяції, а концентрація інших алелей може випадково підвищитися при подальшому спалаху чисельності.
Штучний добір — це вибір і подальше розмноження організмів, що мають господарсько-цінні або декоративні ознаки і властивості (продуктивність, довжина шерсті, кількість молока, розмір плодів, кількість насіння і т. д.), який робить людина. Виведені сорти рослин або породи тварин несуть один або декілька гіпертрофованих ознак, зумовлених надмірною активацією певних генів. Такі організми не можуть вижити в дикій природі. Ч. Дарвін показав, що між природним і штучним добором сполучною ланкою є несвідомий добір як основна форма відбору, виконаного людиною на перших щаблях її культурного розвитку.
Основні наукові роботи
Ранні роботи (до «Походження видів»)
Незабаром після повернення з подорожі Дарвін видав книгу, відому під скороченою назвою «Подорож натураліста навколо світу на кораблі „Бігль“» (1839). Вона мала великий успіх, і друге, розширене видання (1845) було перекладено багатьма європейськими мовами та безліч разів перевидавалося. Дарвін взяв також участь в написанні п'ятитомної монографії «Екологія подорожі» (1842). Як зоолог Дарвін вибрав об'єктом свого вивчення вусоногих раків, і незабаром став кращим у світі фахівцем з цієї групи. Він написав і видав чотиритомну монографію «Вусоногі раки» (Monograph on the Cirripedia, 1851–1854), якою зоологи користуються до сих пір.
Праця «Походження видів» Ця стаття містить текст, що не відповідає енциклопедичному стилю. Конкретні зауваження можна знайти на сторінці обговорення. Будь ласка, при можливості покращіть цю статтю. (серпень 2012)
В середині липня 1837 Дарвін почав свій записник «B» про Трансмутацію Видів, на сторінці 36 він написав «Я думаю» про своє перше еволюційне дерево
24 листопада 1859 р. вийшла в світ геніальна праця Чарлза Дарвіна «Походження видів шляхом природного добору або збереження обраних рас у боротьбі за життя». Ця книга, в якій так майстерно викладені і всебічно обґрунтовані наукові основи еволюційної теорії, користувалась великою популярністю, і весь її наклад був розпроданий в перший же день. Появу «Походження видів» один із сучасників Дарвіна образно порівнював з вибухом, «якого ще не бачила наука, який так довго підготовлювався і так раптово нагрянув, так нечутно підведений і так смертоносно разючий. За розмірами і значенням заподіяного руйнування, за тим відлунням, яке відгукнулося в найвіддаленіших галузях людської думки, це був науковий подвиг, що не мав собі подібного». Епохальна праця Дарвіна сім разів перевидавалась за життя автора, вона швидко стала відомою науковцям інших країн і була перекладена на більшість європейських мов.
Після публікації «Походження видів» Чарлз Дарвін продовжує енергійно працювати над обґрунтуванням еволюційної теорії. В 1868 р. він публікує капітальну працю «Зміна свійських тварин і культурних рослин», де всебічно аналізує закономірності мінливості, спадковості, штучного добору. Ідею історичного розвитку рослин і тварин Дарвін поширює і на проблему походження людини. В 1871 р. виходить його спеціальна книга «Походження людини і статевий добір», у якій детально аналізуються численні докази тваринного походження людини. «Походження видів» і наступні дві книги становлять єдину наукову трилогію, в яких наведено докази історичного розвитку органічного світу, встановлено рушійні сили еволюції, визначено шляхи еволюційних перетворень, нарешті, показано, як і з яких позицій слід вивчати складні явища і процеси природи. Дарвін опублікував 12 томів своїх творів. Дуже цікавою є його автобіографія «Спогади про розвиток мого розуму і характеру».
Чарлз Дарвін відзначався гострою спостережливістю, досить розвинутими аналітичними і синтетичними здібностями, науковою добропорядністю, винятковою працьовитістю, спрямованістю і акуратністю в роботі. До останніх днів свого життя він не припиняв систематичних наукових досліджень. Так, ще 17 березня 1882 р. Дарвін вів спостереження в своєму саду, а 19 квітня перестало битися велике серце титана людської думки. Похований Дарвін у Вестмінстерському абатстві в Лондоні поряд з Ньютоном, Фарадеєм та іншими видатними науковцями Англії.
Пізні роботи (після «Походження видів»)
1868 року Дарвін опублікував свою другу працю, пов'язану з теорією еволюції — «Зміна тварин і рослин в домашньому стані» (англ. The Variation of Animals and Plants under Domestication), в яку увійшло безліч прикладів еволюції організмів. 1871 року з'явилася ще одна важлива праця Дарвіна — «Походження людини і статевий відбір» (англ. The Descent of Man, and Selection in Relation to Sex), де Дарвін привів аргументи на користь природного походження людини від тварин (мавпоподібних предків). Серед інших відомих пізніх робіт Дарвіна — «Запилення у орхідних» (англ. The Fertilization of Orchids, 1862), «Вираження емоцій у людини і тварин» (англ. The Expression of the Emotions in Man and Animals, 1872); «Дія перехресного запилення та самозапилення в рослинному світі» (англ. The Effects of Cross-and Self-Fertilization in the Vegetable Kingdom, 1876).

