




Основными источниками фолатов являются свежие овощи и зелень: салат, шпинат, капуста, морковь, помидоры, лук.
Фолаты всасываются в проксимальных отделах тонкого кишечника. Всасывание фолатов осуществляются с помощью специфического механизма активного транспорта, требует затраты энергии и обеспечивает поступление фолиевой кислоты в кровоток против концентрационного градиента.
Транспорт фолатов в плазме крови происходит с участием специфических белков, из которых основным является фолатсвязывающий белок из фракции трансферина.
Всосавшиеся фолаты поступают в печень, где накапливаются и превращаются в активные формы. Способность печени накапливать и использовать фолаты находится в зависимости от обеспеченности организма белками, аминокислотами и витаминами. Дефицит витамина В12 и метионина снижает содержание фолатов в печени. Недостаток биотина нарушает образование активной формы витамина – тетрагидрофолиевой кислоты.
Печень играет важную роль в обмене фолатов. При циррозах, жировой инфильтрации печени нарушается накопление и использование фолатов.
Фолиевая кислота образует в организме несколько коферментных форм. Первая стадия образования коферментных форм – это восстановление фолиевой кислоты в тетрагидрофолиевую кислоту при участии дегидрофолатредуктазы. Тетрагидрофолиевая кислота приносит одноуглеродные фрагменты: СН3 – метильная группа, СН2 – метиленовая группа, СН – метиновая, СНО – формильная.
Тетрагидрофолиевая кислота образует следующие коферментные формы:
1. Метилтетрагидрофолиевую кислоту ТГФК-СН3
2. Метилен ТГФК-СН2
3. Метинтетрагидрофолиевую кислоту ТГФК-СН
4. Гидроксиметилентетрагидрофолиевую кислоту ТГФК-СН2ОН
5. Формилтетрагидрофолиевую кислоту ТГФК-СНО
Источником одноуглеродных фрагментов, акцептируемых ТГФК в реакциях катаболизма, являются 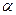 - углеродный атом глицина,
- углеродный атом глицина, 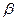 углеродный атом серина, углеродный атом гистидина, а также муравьиная кислота, формальдегид.
углеродный атом серина, углеродный атом гистидина, а также муравьиная кислота, формальдегид.
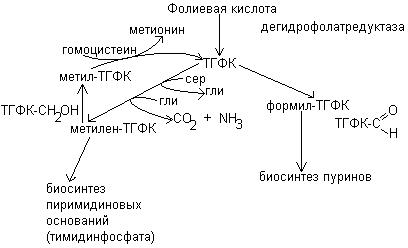
Наиболее важной функцией коферментных форм фолиевой кислоты является их участие в биосинтезе пуриновых оснований и тимидинмонофосфата.
При синтезе пуриновых оснований производные ТГФК служат источником 2 и 8 углеродных атомов пуринового кольца.
Участие коферментных форм фолиевой кислоты в биосинтезе тимидинфосфата, а также пуриновых оснований, входящих в ДНК и РНК, определяют важную роль фолиевой кислоты в синтезе нуклеиновых кислот и процессе пролиферации.
Нарушения обмена фолиевой кислоты.
Нарушения обмена фолиевой кислоты могут быть вызваны следующими причинами:
1. Недостаточное поступление фолатов с пищей.
2. Нарушение всасывания фолатов в организме.
Недостаток фолатов приводит к возникновению анемии. Непосредственным биохимическим дефектом, вызывающим анемию, является торможение биосинтеза ДНК и пролиферации кроветворных клеток, т.к. нарушается синтез пуриновых оснований и тимидинфосфата. Фолатдефицитная мегалобластическая анемия характеризуется снижением количества эритроцитов, гемоглобина и появлением в периферической крови и костном мозге мегалобластов. Введение фолиевой кислоты вызывают быструю гематологическую ремиссию и восстанавливает нормальное кроветворение.
| Известны врожденные дефекты, затрагивающие различные ферменты, участвующие в образовании и взаимопревращениях коферментных форм фолиевой кислоты. |
Известен дефект фолатредуктазы, который приводит к развитию мегалобластической анемии, но эта анемия вызвана нарушением превращения в формил-ТГФК.
Витамин В12-кобаламин.
Группа витаминов В12. содержит атом кобальта, соединенный с 4 восстановленными пиррольными кольцами.
Синтезируется исключительно микроорганизмами, от них попадает в животные ткани, печень почки. Частично синтезируется микрофлорой кишечника.
Витамин В12 (кобаламин) присутствует в организме в 3 формах: оксикобаламина (НО-В12), метилкобаламина (СН3-В12) и 5-дезоксиаденозилкобаламина (ДАВ12). В составе витамина В12 содержится кобальт. Оксимелиткобаламин является основной транспортной и депонируемой формой В12. метилкобаламин и дезоксиаденозилкобаламин – коферментные формы витамина.
Для всасывания витамина В12 необходим внутренний фактор Кастла – гликопротеид, продуцируемый обкладочными клетками желудка. При резекции желудка нарушается всасывания витамина В12.
После всасывания, витамин В12 связывается со специфическим транспортным белком транскобаламином. Транскобаламин существует в формах I и II. Транскобаламин I транспортирует витамин от кишечника к печени и обеспечивает его депонирование. Транскобаламин II является основной транспортной формой в кровотоке.
Витамин В12 принимает участие в 2 ферментативных реакциях.
1. В виде метилкобаламина катализирует превращение гомоцистеина в метионин путем переноса метильного остатка от метил-ТГФК:
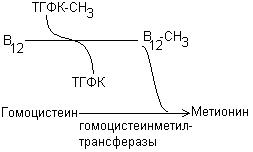
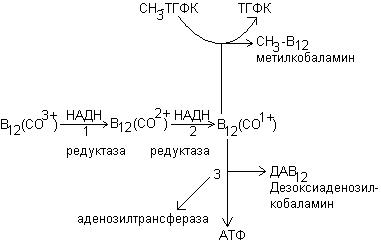
2. Коферментная форма витамина В12-дезоксиаденозилкобаламин необходима для функционирования метилмалонил-КоА-мутазы, которая обеспечивает изомеризацию метилмалонил-КоА в сукцинил-КоА:

| С разветвленной цепью |
 Жирные кислоты Жирные кислоты
| |
| С нечетным числом атомов С |
Холестерин
Изолейцин
Метионин
Треонин
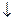
Нарушения обмена витамина В12.
Основными проявлениями недостаточности витамина В12 являются нарушения процессов кроветворения, приводящие к мегалобластической анемии, лейкопении, а также неврологические симптомы дегенерации задних и боковых столбов спинного мозга.
Патология:
1. Механизм развития анемии (гипотеза) – При недостаточности В12 нарушается использование СН3-ТГФК для ресинтеза метионина и вся фолиевая кислота попадает в «ловушку» и ТГФК превращается в СН3-ТГФК и дальше нарушается образование других коферментных форм фолиевой кислоты, которые необходимы для синтеза пуриновых и пиримидиновых нуклеотидов. Это ведет к нарушению биосинтеза нуклеиновых кислот, пролиферации и созреванию кроветворных клеток.
2. Анемия Аддисона-Бирмера (пернициозная анемия) – обусловлена атрофией слизистой оболочки желудка. Атрофия слизистой оболочки, как правило, носит врожденный характер.
3. Врожденные метилпропионатацидемии.
Носит выраженный семейный характер, затрагивает братьев и сестер одного поколения при отсутствии выраженных симптомов заболевания у родителей.
Метилмалонатацидемия обусловлена нарушением образования дезоксиаденозилкобаламина. Это нарушение приводит к накоплению метилмалонил-КоА. Метилмалонил-КоА ингибирует пируваткарбоксилазу и это нарушает превращение пирувата в оксалоацетат и в результате тормозится глюконеогенез, развивается гипогликемия, усиливается катаболизм липидов и ацидоз.
Метилмалонил-КоА тормозит синтез жирных кислот и включается в синтез вместо малонил-КоА, что приводит к появлению жирных кислот с разветвленной углеродной цепью.
Пропионил-КоА включается в синтез жирных кислот и приводит к образованию жирных кислот с нечетным числом атомов углерода. Нарушается синтез липидов и фосфолипидов, происходит нарушение миелиновой оболочки нервной ткани (демиелинизация).
Аскорбиновая кислота (витамин С).
Всасывание аскорбиновой кислоты происходит особенно интенсивно в тонком кишечнике. Всасывание может нарушаться при кишечных дискензиях, энтеритах.
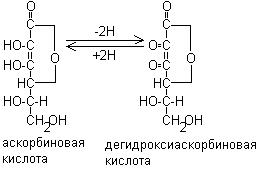
Аскорбиновая кислота, являясь донором водорода, участвует в окислительно-восстановительных реакциях и превращается при этом в дегидроаскорбиновую кислоту:
Аскорбиновая кислота участвует в следующих биохимических процессах:
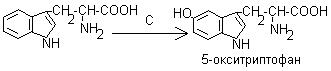
1. Гидроксилирование триптофана в 5-гидрокситриптофан (синтез серотонина).
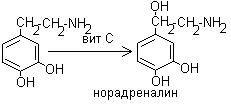
2. Превращение 3,4-дигидрофенилэтиламина в норадреналин
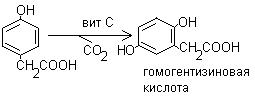
3. Гидроксилирование п-гидроксифенилпирувата в гомогентизиновую кислоту:
4. Гидроксилирование стероидов (циклопентанпергидрофенантрена) при биосинтезе гормонов коры надпочечников из холестерина.
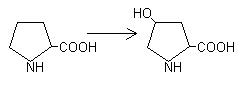
5. Гидроксилирование остатков пролина и лизина при синтезе коллагена:
6. Витамин С принимает участие в работе системы метгемоглобин – гемоглобин.
Аскорбиновая кислота + метгемоглобин  ДАК + гемоглобин
ДАК + гемоглобин
ДАК + глутатион АК + окисленный глутатион
Аскорбиновая кислота восстанавливает метгемоглобин в гемоглобин, сама окисляется в дегидроксиаскорбиновую кислоту. Дегидроксиаскарбиновая кислота восстанавливается под действием глутатиона. Метгемоглобин не накапливается в клетках.
7. Аскорбиновая кислота способствует восстановлению трехвалентного железа в 2-х валентное, которое легче всасывается в кишечнике.
Нарушения обмена витамина С.
Недостаточность аскорбиновой кислоты приводит к цинге. Характерна повышенная ломкость кровеносных капилляров, выявляемая пробой на щипок, болезненность десен, их отечность, разрыхленность, кровоточивость при чистке зубов. В тяжелых случаях изъязвления десен, появление серых налетов, расшатывание и выпадение зубов. Но по мере развития скорбута обнаруживается красновато-синеватые, бурые пятна на голенях, на поверхности бедер.
Витамин А.
Источником витамина А являются продукты животного происхождения: печень трески, морского окуня, сметана, яйца. В моркови, томатах, свекле содержатся каротиноиды, которые являются провитаминами А.
Название «витамин А» объединяет группу соединений: ретинол-спирт, ретинал-альдегид, ретиноевую кислоту. В тканях организма образуются сложные эфиры ретинола с пальмитиновой и уксусной кислотой: ретинилпальмитат, ретинилацетат.
Известны три провитамина А - , , и  -каротины, которые отличаются по строению и биологической активности. Наиболее активен - каротин, который при расщеплении дает 2 молекулы ретиналя. и -каротины дают по 1 молекуле витамина А.
-каротины, которые отличаются по строению и биологической активности. Наиболее активен - каротин, который при расщеплении дает 2 молекулы ретиналя. и -каротины дают по 1 молекуле витамина А.
Для всасывания витамина А необходимы желчные кислоты. В слизистой кишечника ретинол образует эфиры с жирными кислотами и трансформируется в составе хиломикронов. В плазме ретинол связывается ретинолсвязывающим белком и доставляется в ткани. В печени эфиры ретинола депонируются. Часть ретинола в печени окисляется в ретиноевую кислоту, которая выводится с желчью в виде глюкуронидов.
Функции витамина А.
1. Регулирует рост и дифференцировку клеток развивающегося организма. Регулирует дифференцировку и деление быстро делящихся клеток (пролиферирующих) тканей – хряща и костной ткани, эпителия кожи и слизистых.
2. Участие в фотохимическом акте зрения.
3. В форме ретинолфосфата присоединяет глю, фруктозу и транспортирует в клеточную мембрану для синтеза гликопротеидов, т.е. участвует в построении клеточной мембраны (рецепция, контакт, иммунологическое взаимодействие) проницаемость клеточных и субклеточных мембран.
Наиболее изучено участие витамина А в зрительном акте. В этом процессе витамин А участвует в форме цис-ретиналя, который входит в состав светочувствительных пигментов сетчатки глаза. В сетчатке имеется 2 типа клеток – палочки и колбочки. Палочки реагируют на слабое освещение (сумерочное, ночное зрение), а колбочки – на хорошее освещение (дневное зрение) и обеспечивают цветовое зрение. Палочки содержат белок родопсин, а колбочки – йодопсин.
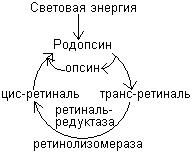
Кванты света поглощаются родопсином и вызывают изомеризацию цис-ретиналя в транс-ретиналь. После чего происходит диссоциация транс-ретиналя и опсина и пигмент обесцвечивается. Изомеризация ретиналя приводит к возникновению электрического импульса, распространяющегося по нервному волокну. В темноте происходит превращение транс-ретиналя в транс-ретинол, а затем в цис-ретинол, который вновь соединяется с белком опсином. Отсутствие регенерации родопсина приводит к слепоте в ночное время или в сумерках.
Нарушения обмена витамина А.
1. Ранним признаком недостаточности витамина А является нарушение темновой адаптации и ночная слепота.
2. Недостаточность витамина А приводит к возникновению фолликулярного гиперкератоза (избыточное ороговение кожи, вызванное задержкой смены эпителия).
3. Ксерофтальмия – сухость конъюктивы глаза.
4. Кератомаляция – размягчение роговицы.
Гипервитаминоз А – возникает довольно быстро.
Причины:
1. Употребление продуктов, богатых витамином А (печень полярных млекопитающих, рыбий жир)
2. Передозировка лечебных препаратов витамина А.
Токсическими дозами являются (острое отравление) 1-6 млн МЕ (но есть индивидуальная чувствительность и тогда меньшая доза может быть токсичной).
Хроническая интоксикация может быть вызвана длительным применением 20 тыс МЕ. Особенно чувствительны дети.
Общие явления: раздражимость, бессонница, головные боли, повышение t, изменения со стороны волос и кожи, боли в суставах, растройство походки, припухлость мягких тканей, увеличение печени, селезенки.
Врожденные нарушения обмена витамина А.
Врожденное нарушение превращения -каротинов в ретиналь.
Дефект фермента, превращающего в слизистой кишечника -каротин в витамин А.
Витамин D.
Витамин D – групповое обозначение нескольких веществ стероидной природы. Наиболее активны эргокальциферол (D2), ходекальциферол (D3) и дигидроэргокальциферол (D4). Образование витамина D3 происходит из холестерина в коже человека при действии ультрафиолетового облучения. Ни один из витаминов не применяется в таких количествах, особенно у детей до 1 года. Потребность ребенка в витамине D – 10 мкг – 400 МЕ в сутки.
Всасывание кальциферолов происходит с помощью желчных кислот. В составе хиломикронов кальциферолы поступают в кровь, а затем в печень. В печени эргокальциферол и холекальциферол гидроксилируются в 25 положении и образуются 25-гидроксихолекальциферол и 25-гидроксиэргокальциферол. После этого поступают в почки, где образуются 1,25-дигидроксикальциферолы.
Функции витамина D.
1. Транспорт ионов кальция и фосфата через эпителий слизистой тонкого кишечника при всасывании.
2. Мобилизация кальция из костной ткани.
3. Реабсорбция кальция и фосфора в почечных канальцах.
В результате действия витамина D повышается содержание ионов кальция и фосфатов в крови.
Витамин D рассматривается как прогормон, т.к. основная функция – обеспечение нормальной концентрации Са в крови (2,5 мМ, 10мг% - очень стабильный, жесткий показатель). Уменьшение содержания Са2+ вызывает судорожное состояние, повышение концентрации Са2+ может вызывать внекостную кальцификацию.
Механизм действия витамина D.
1. Обеспечение всасывания кальция из кишечника (всасывается в виде фосфатов).
2. Реабсорбция кальция в почках.
3. Мобилизация из депо (костная ткань).
Схема обмена витамина D
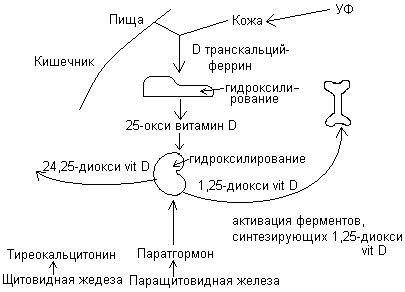
Функции 1,25-диокси витамина D:
1. Деминерализация костей
2. Усиление синтеза белков, связывающих кальций (кальцийсвязывающий белок). Реализует свое действие через генетический аппарат.
3. Усиливает реабсорбцию кальция в почках.
Функции 24,25-диокси витамин D
Усиливает синтез щелочной фосфотазы и Са2+-зависимой АТФ-азы.
Снижение [Са2+] – выброс паратгормона 1,25-диокси витамин D.
Повышение [Са2+] – понижение содержания паратгормона переключение на синтез 24,25-диокси витамин D, выброс тиреокальцитонина усиление снижения кальция в костях.
Нарушение обмена витамина D.
1. Гипервитаминоз по количеству и тяжести на I месте. Гипервитаминоз D возникает при избыточном приеме витамина. Наблюдается деминерализация костей и их переломы. Уровень кальция и фосфатов в крови повышается (они извлекаются из костей, всасываются из кишечника и реабсорбируются в почках). Развивается гиперкальцемия карциноз внутренних органов: аорты, сердца, печени, легких, почек тяжелейшие осложнения, часто смерть.
2. Нарушения обмена витамина D могут возникнуть при заболеваниях почек и печени, где происходит образование активных форм витамина D.
3. Рахит – вызывает недостаток витамина D в пище. При рахите заторможены всасывание ионов кальция и фосфатов, реабсорбция их в почках. Уровень кальция и фосфора в крови снижается, нарушается минерализация костей. Наблюдается размягчение костей, возникают деформации костей конечностей, черепа (замедление зарастания родничков, нарушение челюстно-лицевого скелета), грудной клетки, дряблые мышцы.
Профилактика.
Витамин D назначают не раньше, чем с 1-1,5 месяцев, физиологическая доза 400 МЕ.
Биохимически не обоснованы схемы профилактики, когда витамин D вводят в 1 раз в 1 или 2 недели (всю дозу 3-6-8 тыс. МЕ) и недопустима ударная профилактика рахита, когда вводится доза больше физиологической в 1000 раз.
Врожденные нарушения обмена витамина D.
1. Синдром Де-Тони-Дебре-Факкони.
Характерна триада: гиперфосфатемия, гиперглюкозурия, гипераминоацедемия. В основе генетически обусловленные нарушения транспорта в почечных канальцах. Развивается вторичный рахит (вследствие потери фосфата и ацидоза).
2. Семейный гипофосфатемический витамин D – резистентный рахит.
Гипофосфатемия и признаки рахита на 1-2 году жизни, не поддается лечению витамином D, низкий рост 150-160 см, костные деформации, псевдоатлет (приземистое телосложение).
Причина: снижение реабсорбции неорганического фосфата в почечных канальцах, понижение всасывания в тонком кишечнике. В основе генетические нарушения в третичной структуре белков в почечных канальцах.
Лечение: фосфаты и соли кальция.
3. Врожденный псевдодефицитный витамин D – зависимый рахит – по клинике сходен с рахитом, но отличается резистентностью к стандартным профилактическим и лечебным дозам витамина D.
Характеризуется гипокальциемией и гипераминоацидурией, нет фосфата.
В основе дефект фермента в почках, обеспечивающего образования 1,25-диокси витамин D2/D3.
Витамин К.
Витамин К является хиноном с боковой изопреноидной цепью. Есть два ряда нафтохинонов – филлохиноны и менохиноны. Разработаны синтетические препараты витамина К – менадион, викасол и синкавит.
Витамин К содержится в основном в растительной пище: шпинат, тыква, капуста, томаты, крапива. Частично синтезируется микрофлорой кишечника.
Всасывается витамин К в тонком кишечнике в присутсвии желчных кислот. Транспорт витамина К происходит с хиломикронами. В плазме крова витамин К связывается с альбумином.
Функции витамина К.
1. Витамин К участвует в образовании факторов свертывающей системы крови: II – протромбина, VII – проконвертина, IX – фактора Кристмаса и X – фактора Стюарта.
2. Витамин К в виде кофермента участвует в микросомальном - карбоксилировании глутаминовой кислоты (печень), входящей в состав белков, связывающего кальций.
Протромбин (содержит 10 остатков глу, в отличие от тромбина).
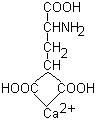
Чтобы протромбин мог активироваться и превратиться в тромбин, он должен связать ионы Са2+. При недостатке витамина К в организме синтезируются дефектные молекулы протромбина, не способные правильно связывать ионы Са2+.
В костях белок – остеокальцин связывает Са2+.
Витамин Е (токоферолы).
Витамин Е представляет группу соединений: , и -токоферолы. Наибольшее значение имеет -токоферол.
Термин «токоферол» происходит от греческого слова «tokos», что в переводе означает «рождение ребенка».
Источником токоферола служат растительные масла.
Для всасывания витамина Е необходимы желчные кислоты. Всасывание происходит путем простой диффузии, транспортируются в составе хиломикронов через лимфатические пути в органы и ткани. В клетках токоферол включается в состав мембран.
Функции витамина Е.
1. Природный антиоксидант – препятствует развитию цепных реакций пероксидного окисления ненасыщенных липидов в биологических мембранах.
2. Токоферол повышает биологическую активность витамина А, т.к. защищает боковую цепь от пероксидного окисления.
Нарушения обмена витамина Е.
Гиповитаминоз Е у человека практически не встречается. Недостаточность витамина Е у экспериментальных животных вызывает атрофию семенников и рассасывание плода, мышечную дистрофию.
При недостатке витамина Е усиливается перекисное расщепления ненасыщенных липидов клеточных мембран, что вызывает выход лизосомальных ферментов.
При Е-авитаминозе повышается склонность эритроцитов к перекисному гемолизу, что приводит к возникновению анемии.
| Витамин | Коферментная (или активная) форма | Тип катализируемой реакции или функция |
| Биотин | Биотин | Перенос СО2 |
| АК | Не известна | Кофактор реакций гидроксилирования |
| Вит А | Ретиналь | Зрительный процесс |
| Вит Е | Не известна | Защита мембранных липидов |
| Вит D | 1,25-дигидрокси-холекальциферол | Регуляция обмена Са2+ |
| Вит К | Не известна | Кофактор реакций карбоксилирования. |

