




Приток крови к мышце и, следовательно, снабжение ее кислородом зависит от работы, которую она совершает. Количество кислорода, которое необходимо доставить к работающей мышце, в 500 раз превышает потребность в кислороде мышцы, находящейся в состоянии покоя. Поэтому при мышечной работе количество крови, поступающее в мышцу, сильно возрастает (300–500 капилляров/мм3 объема мышцы) и может в 20 раз превышать этот показатель для неработающей мышцы.
Даже в состоянии покоя мышцы всегда находятся в активном (непроизвольном) напряжении (тонусе). Тонус скелетных мышц поддерживается за счет постоянно поступающих в них импульсов (рефлексный тонус). Эти импульсы не ощущаются, поскольку двигательные элементы мышечных волокон также стимулируются. Мышечный тонус контролируется самостоятельно посредством мышечного веретена и сухожилий. При отсутствии мышечного тонуса говорят о вялом (атоническом) параличе. При усиленном тонусе развивается спастический паралич.
Если мышца в течение долгого времени не выполняет работу или нарушается ее иннервация, то она атрофируется. С другой стороны, при повышенной нагрузке на мышцы, например у спортсменов, происходит утолщение отдельных мышечных волокон и наступает гипертрофия мышц. При сильных повреждениях мышцы формируется шрам из соединительной ткани, поскольку способность мышц к регенерации ограничена.
Боль, возникающая в мышце при непривычной или усиленной нагрузке, свидетельствует о ее чувствительности. Существует мнение, что это связано с накоплением лактата, который образуется при анаэробном (недостаток кислорода) распаде глюкозы или других продуктов, либо является результатом мелких повреждений в самих мышцах (микроповреждений). Боль развивается не только в нетренированной мышце, но также после оперативных вмешательств или при мышечных судорогах.
Мышечная судорога – в основном обратимое самопроизвольное сокращение мышцы, вызванное ее напряжением и происходящее без участия нервного импульса. Это может объясняться или локальной деполяризацией мышечных клеток, связанной с увеличением концентрации ионов К+ в межклеточной среде или высвобождением внутриклеточного Са2+. Судороги, сопровождающиеся развитием чувства усталости, часто связаны со снижением в клетках содержания АТФ из-за дефицита кислорода или глюкозы. Необратимые сокращения мышц называются спазмом.
Некоторое время после смерти мышцы остаются мягкими вследствие прекращения тонического влияния мотонейронов. Через 4–10 ч после смерти в мышцах прекращаются обменные процессы, и поэтому исчерпываются запасы АТФ и возможность разъединения головки миозина с актиновым филаментом исчезает. Возникает явление трупного окоченения с выраженной ригидностью скелетных мышц. Как правило, процесс начинается с мышц нижней челюсти. Через 1-3 дня окоченение мышц проходит. К этому времени начинает разрушаться структура тканей (автолиз).
Мышечная ткань сердца представлена специфической формой поперечнополосатых мышц (рис. 1.25).По сравнению со скелетными мышцами, она имеет следующие особенности:
1. В отличие от краевого расположения ядер в клетке скелетных мышц, ядра в клетке мышечной ткани сердца располагаются в центре клетки. Сами клетки по диаметру меньше мышечных волокон скелетных мышц.
2. В противоположность мышечным волокнам скелетных мышц, снаружи не имеющих фибриллярных структур, необходимых для связывания между собой, клетки мышечной ткани сердца связаны друг с другом вставочными дисками. Такая организация мышечных клеток сердца дает возможность возникающему в синусном узле импульсу веерообразно распространяться по стенкам обоих предсердий, и далее по внутренней поверхности желудочков.
3. Еще одна особенность сердечной мышцы заключается в способности некоторых ее клеток генерировать импульсы не только в ответ на внешние раздражители, но и спонтанно. Вместе с тем активность клеток мышцы сердца находится под контролем автономной нервной системы. Например, по сигналу симпатической нервной системы скорость сокращений сердца может усиливаться, а по сигналу парасимпатической – замедляться.
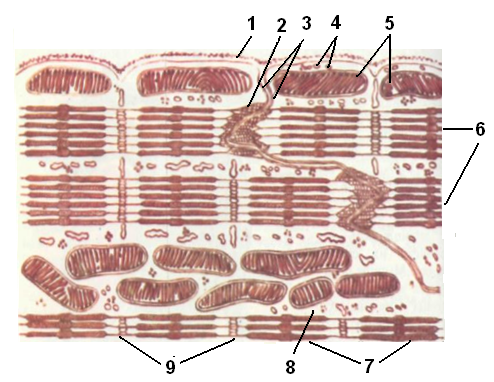
Рис. 1.25. Схема строения кардиомиоцита:
1 – базальная оболочка мышечного волокна, 2 – окончание миопротофибрилл на цитолемме кардиомиоцита, 3 – вставочный диск между кардиомиоцитами, 4 – саркоплазматическая сеть, 5 – саркосомы (митохондрии), 6 – миопротофибриллы, 7 – диск А (анизотропный диск), 8 – саркоплазма, 9 – диск И (изотропный диск)
Преобладающая часть сердечной стенки – миокард, т. е. мышечный слой. Поперечно-полосатая сердечная мышечная ткань (миокард), отличающаяся от скелетных мышц по строению и функции, состоит из сердечных миоцитов (кардиомиоцитов), образующих соединяющиеся друг с другом комплексы почти прямоугольной формы.
Кардиомиоциты имеют 1–2 овальных ядра, лежащих в центре; миофибриллы расположены по периферии строго прямолинейно. Характерны контакты двух соседних кардиомиоцитов в виде темных полосок, вставочных дисков, которые активно участвуют в передаче возбуждения от клетки к клетке. С помощью дисков кардиомиоциты соединяются друг с другом.
По своему микроскопическому строению сердечная мышечная ткань похожа на скелетную (поперечно-полосатая исчерченность), однако сокращения сердечной мышцы не подконтрольны сознанию человека.
Нервная ткань
Нервная ткань образует центральную нервную систему (ЦНС) – головной и спинной мозг и периферическую (ПНС) – нервы с их концевыми пластинками, нервные узлы (ганглии). Нервная ткань обеспечивает анализ и синтез сигналов (импульсов), поступающих в мозг. Она устанавливает взаимосвязь организма с внешней средой и участвует в координации функции внутри организма, обеспечивая его целостность (вместе с гуморальной системой – кровью, лимфой).
Нервную систему условно подразделяют на две части: соматическую (анимальную) и вегетативную (автономную). Соматическая нервная система иннервирует главным образом тело, поперечнополосатые, или скелетные мышцы, кожу, обеспечивает связь организма с внешней средой. Вегетативная (автономная) нервная система иннервирует все внутренности, железы, в том числе и эндокринные, гладкие мышцы органов и кожи, сосуды и сердце, а также обеспечивает обменные процессы во всех органах и тканях.
Нервная ткань состоит из нервных клеток – нейронов (нейроцитов), отличающихся особым строением и функцией, и нейроглии, которая выполняет опорную, трофическую, защитную и разграничительную функции.
Нервная клетка (нейрон) имеет тело и отростки различной длины, является морфофункциональной единицей нервной системы. Нейрон состоит из тела с ядром, особых включений, нескольких коротких древовидно ветвящихся отростков и одного длинного отростка. Нервные клетки способны воспринимать раздражения из внешней или внутренней среды, трансформировать (преобразовывать) энергию раздражения в нервный импульс, проводить их, анализировать и интегрировать. Нервная клетка динамически поляризована, т. е. способна пропускать нервный импульс только в одном направлении. Длинный отросток, по которому нервный импульс движется от тела нервной клетки к концевым аппаратам, к рабочим органам (мышце, железе) или к другой нервной клетке, называется аксоном (нейритом). Другие, более короткие отростки, по которым нервный импульс направляется к телу клетки, называются дендритами.
В зависимости от количества отростков различают униполярные (одноотростчатые), биполярные (двухотростчатые) и мультиполярные (многоотростчатые) нервные клетки (рис. 1.26).К биполярным относятся и ложноуниполярные нейроны – рецепторные нейроны спинномозговых ганглиев, у которых проксимальные отделы отростков сливаются между собой, а затем вскоре Т-образно делятся на аксон и дендрит. Размеры тел нервных клеток колеблются в пределах от 4–5 до 130–140 мкм, а длина отростков может достигать метра и более.
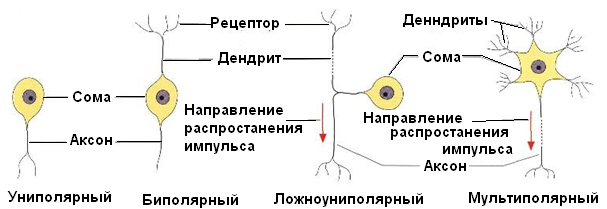
Рис. 1.26. Типы нейронов
Основная особенность нейронов – наличие телец Ниссля и нейрофибрилл (рис. 1.27), которые необходимы для синтеза нейромедиаторов и ферментов. Тельца Ниссля представляют собой собранный в агрегаты гранулярный эндоплазматический ретикулум с полирибосомами и большим количеством мРНК. Нейрофибриллы образованы микротрубочками и нейрофиламентами. Через нейротрубочки осуществляется транспорт нерастворимых белков (ферментов, медиаторов) в синапс. В аксонах отсутствуют элементы эндоплазматической сети и комплекса Гольджи. В дендритах имеются элементы зернистой эндоплазматической сети и рибосомы.
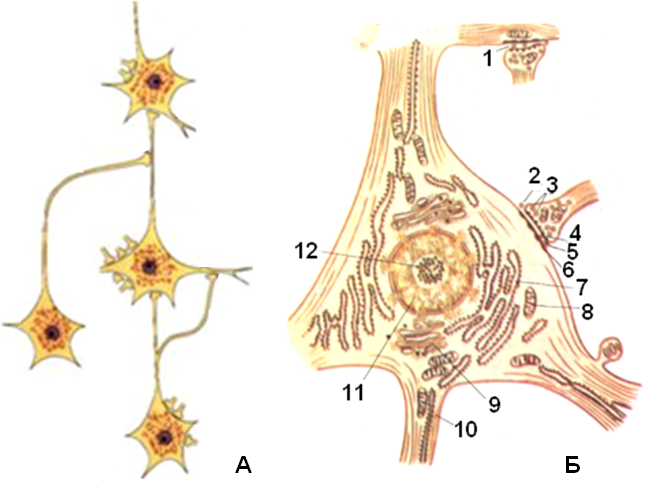
Рис. 1.27. Схема устройства цепочек нейронов и строения нервной клетки:
А – различные типы синапсов, Б – схема ультрамикроскопического строения нервной клетки: 1 – аксонодендритический синапс, 2 – аксоносоматический синапс, 3 – пресинаптические пузырьки, 4 – пресинаптическая мембрана, 5 – синаптическая щель, 6 – постсинаптическая мембрана, 7 – эндоплазматическая сеть, 8 – митохондрия, 9 – внутриклеточный сетчатый аппарат, 10 – нейрофибриллы, 11 – ядро, 12 – ядрышко
Нейроны в нервной системе образуют цепочки, которые передают возбуждение от точки восприятия раздражения в центральную нервную систему и далее к рабочему органу. Нейроны связаны между собой с помощью отростков, которые образуют множество межклеточных контактов – синапсов (от греч. synapsis – связь), передающих нервный импульс от одного нейрона к другому.
Различают синапсы – аксосоматические, когда окончания аксона одного нейрона образуют контакты с телом другого, аксодендритические, когда аксоны вступают в контакт с дендритами, а также аксоаксоналъные и дендродендритические, когда контактируют одноименные отростки, и т. д. Такое устройство цепочек нейронов создает возможность для проведения возбуждения по одной из множества цепочек нейронов благодаря наличию физиологических контактов в определенных синапсах и физиологическому разъединению в других. Количество дендритов может достигать 1000 на одну клетку. Аксон, в единственном числе выходящий из сомы, передает импульс или на дендрит другого аксона через синапс, или непосредственно на двигательную концевую пластинку мышечного волокна. Контакт между аксоном и мышечным волокном называется двигательной концевой пластинкой (нейромышечный контакт).
Кроме нейронов в нервной системе имеются клетки нейроглии (глиоциты), выполняющие многообразные функции: опорную, трофическую, защитную и секреторную. Среди них различают две группы: макроглию (эпендимоциты, олигодендроциты и астроциты) и микроглию (рис. 1.28).
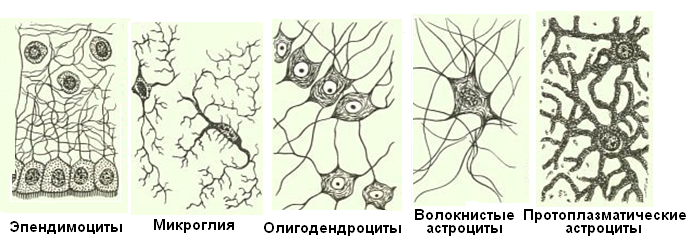
Рис. 1.28. Клетки нейроглии
Клетки нейроглии также существенно различаются по форме, размерам и взаимоотношениям с нейронами. На всем протяжении своего существования клетки нейроглии сохраняют способность к делению. Напротив, нейроны утрачивают эту способность с рождением человека. Поэтому при заболеваниях, сопровождающихся потерей нервных клеток, при различных повреждениях, а также при кислородном голодании глиальные клетки могут замещать нейроны.
Нейроглия окружает нейроны и нервные волокна. Нервные волокна представляют собой отростки нервных клеток, окруженные миелиновыми оболочками. Миелин – многократно закрученный двойной слой плазматической мембраны глиоцита, формирует внутреннюю оболочку осевого цилиндра, наружная образована цитоплазмой и ядром глиоцита (рис. 1.29).
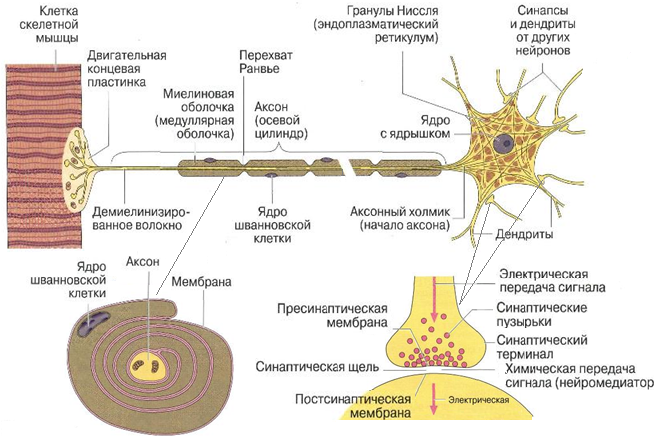
Рис. 1.29. Схема двигательного нейрона, миелиновой оболочки и синапса
Может сформироваться до 100 спиральных слоев миелина правильной пластинчатой структуры. Миелиновые оболочки содержат холестерин, фосфолипиды, некоторые цереброзиды и жирные кислоты, а также белковые вещества, переплетающиеся в виде сети (нейрокератин). Миелиновая оболочка образована особым типом глиальных клеток: шванновскими клетками (в периферической нервной системе) и олигодендроцитами (в центральной нервной системе). Химическая природа миелина периферических нервных волокон и миелина центральной нервной системы несколько различна. Эти два вида миелина обладают и различными антигенными свойствами, что выявляется при инфекционно-аллергической природе заболевания. При формировании миелина ЦНС один олигодендроглиоцит имеет связи с несколькими сегментами миелина нескольких аксонов; при этом к аксону примыкает отросток олигодендроглиоцита, расположенного на некотором расстоянии от аксона, а внешняя поверхность миелина соприкасается с внеклеточным пространством. Шванновская клетка при образовании миелина ПНС формирует спиральные пластинки миелина и отвечает лишь за отдельный участок миелиновой оболочки между перехватами Ранвье. Перехваты соответствуют границе двух смежных глиоцитов. Миелиновая оболочка обеспечивает роль электрического изолятора. Кроме того, предполагается ее участие в процессах обмена осевого цилиндра. Цитоплазма шванновской клетки вытесняется из пространства между спиральными витками и остается только на внутренней и наружной поверхностях миелиновой оболочки. Эта зона, содержащая оттесненную цитоплазму шванновских клеток и их ядра, называется наружным слоем и является периферической зоной нервного волокна.
Различают безмиелиновые и миелиновые волокна. И те и другие волокна снаружи покрыты базальной мембраной. Миелиновые волокна значительно толще безмиелиновых нервных волокон. Диаметр поперечного сечения их колеблется от 1 до 20 мкм. Роль миелиновой оболочки в стимуляции передачи сигнала связана со способностью импульса «перепрыгивать» с одного перехвата на другой (сальтаторное проведение импульса). Имеется прямая зависимость между толщиной этой оболочки и скоростью проведения импульсов. Волокна с толстым слоем миелина проводят импульсы со скоростью 70–140 м/с, в то время как проводники с тонкой миелиновой оболочкой со скоростью около 1 м/с и еще медленнее – безмякотные (безмиелиновые) волокна (0,3–0,5 м/с), т.к. в безмиелиновом волокне проведение нервного импульса (волна деполяризации мембраны) идет не прерываясь по всей плазмолемме.
Способность отвечать возбуждением на внешние сигналы характерна для всех клеток. Быстрая передача сигналов посредством специализированных структур (аксонов) присуща только нервным клеткам. Однако следует отметить, что аксоплазма не служит проводником импульса, хотя является раствором электролитов. Удельное сопротивление аксоплазмы превышает сопротивление меди в 108 раз. Таким образом, электрический сигнал, отвечающий отдельному импульсу, образуется не из-за перемещения ионов вдоль аксона по аксоплазме, а благодаря перемещению ионов в области перехвата Ранвье в перпендикулярном аксону направлении при открытии потенциал-зависимых каналов и работе энергозависимых ионных насосов. Распространение нервного импульса представляет собой самоподдерживающийся процесс, благодаря тому, что при деполяризации мембраны возникают электические токи, замыкающиеся через наружную проводящую среду на соседние участки мембраны (в случае миелинового волокна – на соседние перехваты Ранвье) и отрывающие «ворота» потенциал-зависимых ионных каналов (пор) в мембране аксона (рис. 1.30).

Рис. 1.30. Возникновение замкнутых электрических токов
Первичное возникновение потенциала действия зависит от кратковременной деполяризации мембраны или от изменения ее заряда, который, в свою очередь, распространяется по всему нейрону, включая аксон. Деполяризация мембраны наступает под действием импульса, полученного нервной клеткой от сенсорной клетки (в результате действия внешнего сигнала: свет, механическое, термическое или болевое раздражение) или от другой нервной клетки при синаптической передаче с участием конкретных химических соединений (нейромедиаторов).
Для нервной системы животных и человека сигнал, или потенциал действия, представляет собой универсальное средство сообщения. Существенным параметром такой связи является не интенсивность одиночного потенциала действия, а количество полученных, обработанных и переданных нервным волокном сигналов в единицу времени (частота). Таким образом, язык, или код нейрона, выражается частотой сигнала (до 500 импульсов в секунду).
Для нервной клетки характерен отрицательный потенциала покоя, который выражается разностью электрических потенциалов между наружной клеточной мембраной и содержимым клетки. При возбуждении нервной клетки раздражителями электрической или химической природы происходит кратковременная потеря положительного потенциала на ее мембране, и она заряжается слабо отрицательно. Мембранный потенциал меняется от –60 мВ (потенциал покоя) до +20 мВ. Менее чем за 1 мс исходный потенциал восстанавливается. Поскольку клетка теряет первоначальную поляризацию, этот процесс называется деполяризацией. Возвращение клетки к исходному состоянию носит название реполяризации.
Механизм генерации потенциала действия следующий (рис. 1.31). Когда сигнал достигает нервной клетки, на короткое время приоткрываются мембранные поры (каналы), пропускающие только ионы натрия (Na+). Это позволяет им поступать в клетку из-за существующего градиента концентрации (в клетке содержится мало ионов Na+). В результате в клетке создается избыток положительно заряженных ионови происходит деполяризация мембраны. Однако менее чем через 1 мс Ка+-каналы закрываются и открываются дополнительные К+-каналы. Из клетки начинают выходить ионы К+, что приводит к реполяризации мембраны и к восстановлению исходного потенциала покоя. После закрытия К+-каналов из клетки должны быть удалены ионы натрия (ионный насос). Этот процесс сопровождается затратой энергии (АТФ). После этого клетка готова к следующему акту возбуждения.
Распространение потенциала действия по поверхности нервной клетки или вдоль аксона происходит потому, что при замыкании возникающих электрических полей открываются соседние ионные каналы. Таким образом, возбуждение мембраны распространяется на всю клетку и вдоль аксона. Скорость передачи импульса составляет от нескольких метров в секунду (автономные нервы) до 120 м/с (двигательные нервы произвольных мышц).
Нервный импульс, поступающий в пресинаптическое окончание, вызывает освобождение в синаптическую щель медиатора, который действует на постсинаптическую мембрану, вызывая образование нервного импульса в постсинаптической части. Передача импульса с аксона на другой нейрон происходит через синапс при участии нейромедиаторов. В своей пресинаптической части аксон образует булавовидное утолщение, синаптический терминал, содержащий многочисленные мелкие гранулы, синаптические пузырьки. Когда потенциал действия достигает терминала, находящиеся в пузырьках молекулы медиатора за счет экзоцитоза начинают поступать в синаптическую щель (см. рис. 1.30). Они проходят через синаптическую щель (10–50 нм) за счет диффузии, попадают на постсинаптическую мембрану и связываются с соответствующими рецепторами. При связывании происходит или деполяризация постсинаптической мембраны, способствующая дальнейшему прохождению сигнала (возбудительный синапс), или ее гиперполяризация, которая препятствует его проведению (тормозной синапс). Нейромедиаторы диффундируют через синаптическую щель и вызывают деполяризацию постсинаптической мембраны, способствующую дальнейшей передаче импульса. Синапсы, в которых передача осуществляется с помощью биологически активных веществ, называются химическими. Наряду с химическим имеются электротонические синапсы, в которых передача импульса происходит непосредственно биоэлектрическим путем между контактирующими клетками.
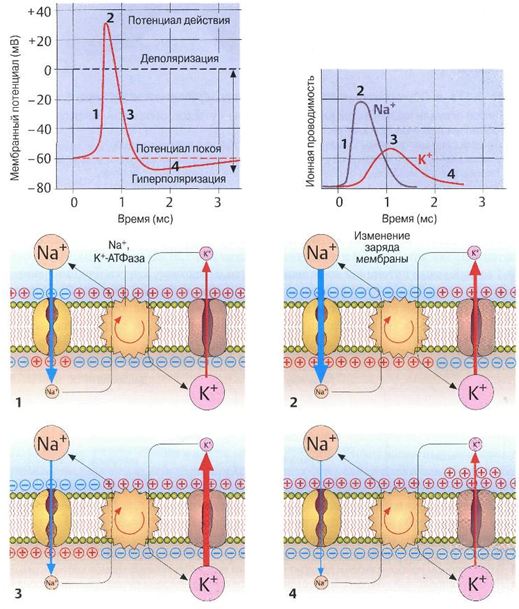
Рис. 1.31. Изменение потенциала действия во времени
К числу медиаторов, передающих сигнал возбуждения, относятся ацетилхолин и глутамат. Передача торможения осуществляется глицином и γ-аминомасляной кислотой (ГАМК). Многие медиаторы проявляют более сложные эффекты, и поэтому их нельзя отнести к какой-то одной группе. К их числу относится норадреналин, дофамин, серотонин и эндогенные опиоиды (эндорфины, энкефалины, динорфины).
Нейромедиаторы относятся к различным классам химических соединений. Норадреналин, серотонин и дофамин представляют собой биогенные амины, глутамат и глицин – аминокислоты, а эндогенные опиоиды являются пептидами (нейропептидами).
Некоторые экзогенные опиоиды, например морфин, выполняют функцию эндогенных нейромедиаторов белковой природы и связываются с такими же мембранными рецепторами. При этом блокируется передача болевых сигналов.
Серотонин представляет собой пример нейромедиатора, который влияет на различные функции мозга. Он участвует в регуляции температурного баланса организма и в формировании ощущений, является стимулятором двигательной активности, влияет на настроение и чувства человека. В основе действия многих психотропных веществ лежит их способность влиять на метаболизм серотонина.
Поскольку импульс всегда проходит в одном направлении, например от аксона, связанного с другой нервной клеткой, синапс можно рассматривать как клапан. Некоторые синапсы способствуют передаче сигнала или тормозят ее. Наконец, синапсы играют важную роль в процессах памяти и обучения. Чем чаще синапс передает какой-либо сигнал, тем быстрее этот сигнал начинает передаваться. На протяжении жизни человека синапсы могут утрачиваться.
Астроциты, мелкие глиальные клетки, участвуют в формировании так называемого гематоэнцефалического барьера (ГЭБ), который препятствует прохождению многих макромолекул, токсинов, лекарств из крови в головной мозг, за счет многочисленных псевдоподий (широких расширяющихся на концах «ножек»), которые, контактируя между собой, со всех сторон окружают стенки капилляров (рис. 1.32).
.
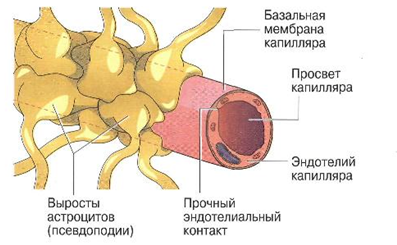
Рис. 1.32. Схема образования гематоэнцефалического барьера
В отличие от других органов, клетки эндотелия капилляров в ЦНС очень плотно примыкают друг к другу. ГЭБ окружает мозг и функционирует как регулятор, позволяющий углеводам (глюкозе) и белкам поступать в мозг только по специфическим транспортным путям. При этом жирорастворимые вещества проходят в мозг почти свободно. Поэтому многие соединения, в том числе большинство лекарств, нерастворимых в жирах, не преодолевают такой барьер. Скопление в спинном и головном мозге тел нейронов и дендритов составляет серое вещество мозга, а отростки нервных клеток образуют белое вещество мозга. Тела нервных клеток формируют скопления и называются ядрами в центральной нервной системе и ганглиями (нервными узлами) – в периферической. По функциям выделяют три вида нейронов:
– чувствительные (афферентные) – проводят нервный импульс от органов в мозг;
– двигательные (эфферентные) –от мозга к органам;
– вставочные (ассоциативные) – располагаются в мозгу, переключают нервный импульс с чувствительных нейронов на двигательные.
В мозжечке и в больших полушариях клетки образуют слоистые (стратифицированные) структуры, называемые корой. Кора покрывает всю поверхность больших полушарий. Ее структурными элементами являются нервные клетки с отходящими от них отростками – аксонами и дендритами – и клетки нейроглии. В коре полушарий большого мозга человека насчитывают около 12–18 млрд нейронов. Основная масса клеток коры состоит из нейронов с пирамидной, веретенообразной и звездчатой формой клеток. Полагают, что пирамидные и веретенообразные клетки с длинными аксонами представляют преимущественно эфферентные системы коры, а звездчатые – преимущественно афферентные. Клеток нейроглии в головном мозге в 10 раз больше, чем ганглиозных (нервных) клеток, около 100–130 млрд. Толщина коры варьирует от 1,5 до 4 мм. Общая поверхность обоих полушарий коры у взрослого человека составляет от 1450 до 1700 см2.
Термин нерв используется только для периферической нервной системы. Нерв состоит из сотен индивидуальных аксонов, заключенных в миелиновые оболочки, а также кровеносных и лимфатических сосудов. В одном нерве могут находиться как чувствительные (афферентные), так и двигательные (эфферентные) волокна. Нерв окружен оболочкой из соединительной ткани.
В отличие от аксонов в ЦНС, периферические нервы способны к регенерации после повреждений, даже если нерв перерезан. Это происходит при сшивании концов нерва. После перерезки нерва, в первую очередь, дегенерирует часть аксона, отделенная от сомы, а шванновские клетки остаются связанными и служат резервом для регенерации аксона. Регенерирующий аксон растет со скоростью 1–2 мм в день в направлении иннервируемого органа (например, мышцы). Для полной реиннервации необходимо несколько месяцев. После ампутации конечности аксоны начинают расти во всех направлениях (так как шванновские клетки также были удалены) и образуют пролиферирующую массу, так называемую ампутационную нейрому.
Органы, аппараты
Орган – анатомически дифференцированная часть организма, состоящая из комплекса тканей, одна из которых выполняет ведущую функцию. Например, печень состоит из всех видов тканей, но основной является эпителиальная (образование желчи и обеспечение обезвреживания веществ, поступающих к печени от органов желудочно-кишечного тракта). «Рабочей» тканью в легких, почках и железах также является эпителиальная ткань, в кости – соединительная (костная), в мозге – нервная ткань. Эпителий покрывает кожу, выстилает слизистые, серозные оболочки и протоки желез. Соединительная ткань выполняет в каждом органе опорную, механическую, трофическую функции, образует соединительно-тканный каркас органа, его строму. Мышечная ткань участвует в образовании стенок кровеносных, лимфатических сосудов, пищеварительной системы, воздухоносных и мочевыводящих путей. Нервная ткань представлена в виде нервов (и их концевых разветвлений), иннервирующих орган, нервных узлов, лежащих в стенках органов.
Орган состоит из структурно-функциональных единиц, отдельных клеток или их популяций, способных выполнять основную функцию органа. Функциональная единица почки – нефрон, печени – печеночная долька.
Органы можно подразделить на внутренние, органы системы опоры и движения и соматосенсорные (органы чувств и кожа). В свою очередь среди внутренних органов различают полые и паренхиматозные. К системе органов опоры и движения принадлежат такие органы, как кости, связки и мышцы.
Все полые органы имеют общий план строения и состоят из трех оболочек: внутренней – слизистой, средней – мышечной и наружной. Наружная оболочка может быть представлена рыхлой соединительной тканью, которая получила название «адвентиция», или серозной оболочкой (брюшина, плевра или перикард). Паренхиматозные органы состоят из стромы – соединительной ткани, образующей каркас, и паренхимы – основного вещества органа.
Таким образом, орган – это относительно обособленное анатомическое образование, структурный элемент, из которого складывается более высокий уровень организации – система органов. В организме органы анатомически и функционально объединяются в системы органов.
Система органов – это интеграция различных органов, имеющих общий план строения, единство происхождения и выполняющих одну большую функцию (например, пищеварения, дыхания).
В организме человека выделяют следующие системы органов: пищеварительную, дыхательную, мочевыделительную, половую, нервную, кровеносную, лимфатическую и иммунную.
Органы, входящие даже в одну систему, существенно различаются по своему строению. Например, в составе пищеварительной системы такие органы, как зубы, язык, пищевод и печень, совершенно различны по своей форме, положению, цвету, консистенции и внутреннему строению. Единство и целостность системы органов определяется, прежде всего, общей направленностью физиологических процессов.
Однако большинство органов в составе одной системы имеют единый план строения. Так, в составе пищеварительной системы глотка, пищевод, желудок, тонкая и толстая кишки имеют общие принципы строения стенки. Они включают слизистую, мышечную и серозную (адвентициальную) оболочки, хотя для каждой из них также характерны типичные особенности строения. Эти органы пищеварительной системы выполняют общую функцию – переваривание пищи и всасывание образовавшихся веществ.
Таким образом, система органов предусматривает, прежде всего, функциональное объединение органов.
Некоторые органы объединяются по функциональному принципу в аппараты.
К аппаратам относят: совокупность органов или систем органов, которые имеют различное строение и происхождение и не связаны анатомически, но выполняют общую функцию (например, опорно-двигательный, эндокринный аппараты); либо совокупность органов или систем органов, которые связаны онтогенетически, но различны по своим функциональным задачам (например, мочеполовой аппарат).

