




Як показує експеримент, для біологічних тканин характерні великі значення кута зсуву фаз між силою струму і напругою, причому сила струму випереджає за фазою напругу. Це свідчить, що частка ємнісного опору в біологічних об’єктах значна. Наведемо деякі значення кута зсуву фаз j при частоті 1 кГц для різних біооб’єктів:
нерв жаби............ – 64°
м’яз кролика....... – 65°
шкіра жаби.......... – 55°
шкіра людини...... – 55°
ламінарія............. – 78°
Як правило, індуктивністю біологічних об’єктів нехтують (при частотах n < 1010 Гц) і вважають, що їхній імпеданс дорівнює геометричній сумі активного R і ємнісного XС опорів. Для характеристики пропускання струму живими клітинами використовують еквівалентні схеми, тобто такі комбінації С і R, які можуть моделювати електричні параметри біологічних тканин. Розглянемо найпростіші з них.

Мал. 4.12.
При послідовному сполученні R і C (мал. 4.12а) схема має суттєві розбіжності з дослідом при пропусканні постійного струму. Якщо w ®0, то вона чинить нескінченно великий опір постійному струмові(Z 0®¥), що не відповідає дійсності. Якщо ж С і R з’єднати паралельно (мал. 4.12б), то при високих частотах (w ® ¥) Z ¥® 0, що теж не відповідає дійсності. Для біологічних тканин характерне більш складне поєднання ємності й активного опору. Найбільш вдалими є схеми, що приведені на мал. 4.12в,г. При пропусканні через такі схеми високочастотного струму при w ® ¥, XC ® 0 їх імпеданс асимптотично наближається до деякої константи Z ¥ (у випадку в: Z ¥ = R 1× R 2/(R 1 + R 2), а у випадку г: Z ¥ = R 1 + R 2). Ці схеми з певним наближенням можуть бути еквівалентними електричними схемами біологічних тканин, але ні одна з них не може повністю відтворювати закономірності пропускання електричного струму через біологічні системи.
На мал. 4.13 приведені графіки дисперсії (частотної залежності) імпедансу рослинної тканини: 1 – крива для нормальної здорової тканини; 2 – для нагрітої до t = 50°C протягом 2 хв; 3 – те ж саме протягом 4 хв; 4 – після кип’ятіння в воді протягом 20 хв.

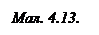 З малюнка видно, що крутизна кривої в міру відмирання тканини зменшується, тобто спостерігається зменшення низькочастотного опору, тоді як при високих частотах імпеданс практично залишається постійним. Причиною є те, що при відмиранні руйнуються мембрани – “живі конденсатори”, і нежива біологічна тканина володіє суто омічним опором. Вказані особливості використовуються для оцінки фізіологічного стану об’єктів. Як правило, розраховується коефіцієнт дисперсії К, який дорівнює відношенню імпедансу при низьких частотах (102 – 104 Гц) до імпедансу при високих (106 – 108 Гц) частотах:
З малюнка видно, що крутизна кривої в міру відмирання тканини зменшується, тобто спостерігається зменшення низькочастотного опору, тоді як при високих частотах імпеданс практично залишається постійним. Причиною є те, що при відмиранні руйнуються мембрани – “живі конденсатори”, і нежива біологічна тканина володіє суто омічним опором. Вказані особливості використовуються для оцінки фізіологічного стану об’єктів. Як правило, розраховується коефіцієнт дисперсії К, який дорівнює відношенню імпедансу при низьких частотах (102 – 104 Гц) до імпедансу при високих (106 – 108 Гц) частотах:
 . (4.9)
. (4.9)
У нормальних тканинах К залежить від положення організму в еволюційному ряді. Наприклад, коефіцієнт дисперсії печінки ссавців дорівнює приблизно 9–10, печінки жаби – 3–4. При відмиранні тканин вказаний коефіцієнт наближається до одиниці. Критерієм життєздатності є також частотна залежність тангенса кута зсуву фаз між силою струму і напругою. Вона дає уявлення про співвідношення між величинами активного і реактивного опорів:
tg j = (XС – XL)/ R» XC / R.
Імпеданс тканин залежить також від їх функціонального стану, і це використовується в діагностиці. Імпеданс кровоносних судин залежить від їх кровонаповнення, а значить і від серцево-судинної діяльності. На цьому базується діагностичний метод, який називають реографією. Реографія вивчає залежність активної складової імпедансу біологічної тканини від її деформації в процесі серцевої діяльності. Отримують реограми серця, головного мозку, магістральних судин, легень тощо. Відповідно до методики Кедрова вважається, що відносна зміна об’єму ділянки кровоносної судини (чи іншої біологічної тканини) D V/V прямо пропорційна зміні опору D R / R:
D V / V ~ D R / R.
Співвідношення між активною та реактивною складовими опору може змінюватись при зміні фізіологічного стану та деяких патологіях. Наприклад, при запаленні на перших стадіях спостерігається збільшення активного опору тканин. Цей ефект обумовлений тим, що струм низької частоти йде переважно через міжклітинну рідину, яка володіє суто омічним опором. При запаленні клітини набухають і переріз міжклітинних ділянок зменшується, що й призводить до підвищення омічного опору. Ємнісний опір при цьому практично не змінюється, оскільки не змінюється структура клітин. Очевидним є той факт, що за незмінного ємнісного опору збільшення омічного опору свідчить про набухання клітин, а зменшення омічного опору, навпаки, свідчить про зменшення об’єму клітин. Зворотний ефект спостерігається на ранніх стадіях онкологічних захворювань. Перетворення нормальних клітин у ракові супроводжується появою молодих клітин і підвищенням ємнісного опору.
5. Проходження змінного електричного струму через біологічні об'єкти.
Кожній клітині, окрім омічного опору, властивий також ємнісний, зумовлений накопиченням іонів протилежного знаку біля клітинних мембран. Поляризаційна ємність сягає 10 мкФ та більше на квадратний сантиметр поверхні мембрани. Наявність електрорушійної сили поляризації та поляризаційної ємності ускладнює вимірювання електропровідності живих клітин при постійному струмові. Окрім цього, цей струм, проходячи через цитоплазму, розкладає її. З цієї причини електричні параметри біологічних об'єктів зручніше вимірювати з використанням змінного струму.
Проходження змінного струму в колі описується законом Ома: І=  ,
,
де  - повний опір кола (імпеданс).
- повний опір кола (імпеданс).
У результаті досліджень з'ясовано:
1. Опір біологічного об'єкта при змінному струмі менший, ніж при постійному.
2. Опір зменшується під час збільшення частоти змінного струму до деякого значення, після чого залишається практично сталим. Це явище називається дисперсією імпедансу. Воно зумовлене залежністю ємнісного опору від частоти, а також поляризаційними процесами, які внаслідок інерції іонів послаблюються за високих частот. Опір більшості тканин мінімальний при частотах змінного струму - 106 Гц, а нерва - 109 Гц.
3. За певної частоти опір біологічного об'єкта залишається сталим, якщо не змінюється його фізіологічний стан. Внаслідок ушкодження тканини опір її зменшується до певного мінімального значення, яке буде відповідати її відмиранню. Дисперсія імпедансу спостерігається лише в живих тканинах. Після відмирання тканини опір від частоти не залежить.
Частотні характеристики електричного опору різних тканин подібні, але значення повного опору (імпедансу) — різні. Наприклад, кісткова тканина містить багато кристалів фосфату кальцію, тому має більший імпеданс, ніж м'які тканини.
Метод вимірювання імпедансу використовують у медичних дослідженнях, а саме: при вивченні процесів у живих клітинах і тканинах за впливу випромінювання, ультразвуку та інших фізичних чинників, а також під час зміни фізіологічного' стану. Наприклад, виявлено, що внаслідок запальних процесів на перших стадіях хвороби збільшується опір тканини. Це пояснюється тим, що під час запалення клітина набрякає, міжклітинні проміжки зменшуються, активний опір збільшується. На наступних стадіях запального процесу змінюються структура та хімічний склад клітин, що призводить до зменшення ємності та опору.
Отже, вимірювання електричних параметрів тканин може використовуватись як засіб діагностики стадій запальних процесів.
Виявлено, що на різних стадіях утворення злоякісних пухлин ємнісний опір тканини змінюється. Тому цей показник можна використати для ранньої діагностики захворювання. Під час відмирання тканини, а також під дією ушкоджувальних чинників (радіація, ультразвук, температура) збільшується проникність мембран і, як наслідок, збільшуються іонні потоки — послаблюється ефект поляризації на межі розділу. Це зумовлює зменшення опору та ємності об'єкта за низьких частот, а за високих — поляризація на межі розділу практично відсутня, тому високочастотний опір майже не змінюється. Таким чином, під час дії ушкоджувальних чинників та відмирання тканини дисперсія її електричних параметрів зменшується. Дисперсія таких параметрів мертвої тканини взагалі відсутня.
Для оцінки життєздатності тканин вводять коефіцієнт поляризації:
К= 
де  опір тканини за низьких частот струму;
опір тканини за низьких частот струму;  опір тканини за високих частот.
опір тканини за високих частот.
Наприклад, для печінки ссавців К= 9-10, а печінки жаби — 2-3. К залежить від інтенсивності обміну речовин у тканинах. Під час відмирання тканин коефіцієнт поляризації зменшується, а для мертвої — наближається до одиниці.
У клінічній практиці вимірювання імпедансу використовують для дослідження кровонаповнення органів. Метод реєстрації зміни імпедансу органів під час їх кровонаповнення називають реографією. Використовуючи багатоканальні реографи, можна досліджувати перерозподіл крові між органами в нормі і патології.
Реоенцефалографія — це метод дослідження мозкового кровообігу, який грунтується на реєстрації пульсових коливань імпедансу головного мозку під час проходження через нього струму високої частоти, але малої сили та напруги. Цим методом визначають стан гемодинаміки, характер пульсового кровонаповнення окремих ділянок головного мозку, стан стінок судин та венозного кровообігу.
У фізіотерапії використовують ультрависокочастотні електричні поля. Ці поля зумовлюють поляризаційні явища у тканинах, і, як наслідок, виникає тепловий ефект, який залежить від діелектричної проникності та опору тканин, частотних характеристик поля. Максимальне нагрівання тканин спостерігається в області дисперсії електропровідності, тобто під час інтенсивної поляризації. Тому важливим напрямом розвитку фізіотерапії є дослідження електричних властивостей тканин у діапазонах частот, що зумовлюють лікувальний ефект під час фізіопроцедур.

