




КАФЕДРА ГИСТОЛОГИИ, ЦИТОЛОГИИ И ЭМБРИОЛОГИИ
МЕТОДИЧЕСКИЕ УКАЗАНИЯ ПО ЭМБРИОЛОГИИ
ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ СТУДЕНТОВ
ОДЕССА - 2008
УДК: 611-013.1 – 611-013.86
Методические указания по эмбриологии для самостоятельной работы студентов (учебное пособие к практическим занятиям по эмбриологии для студентов медицинского и стоматологического факультетов) созданы на основе методических указаний по эмбриологии, изданных коллективом кафедры в 1988 году под редакцией проф. Пчелякова В.Ф. Пересмотрены и дополнены доцентом Арнаутовой Л.В. Одесса, 2008. – 54с.
Настоящее пособие к практическим занятиям по эмбриологии предназначено для студентов всех факультетов медицинских вузов. Методическое пособие включает два основных раздела эмбриологии: сравнительную и частную. В разделе сравнительной эмбриологии отражены как общие закономерности индивидуального развития разных представителей животных организмов, так и частные особенности их индивидуального развития. В разделе частной эмбриологии достаточно подробно освещены вопросы эмбрионального развития органов и систем органов, рассматриваются источники и особенности их развития на различных этапах эмбриогенеза, конкретизируется и обобщается предусмотренный программой материал по курсу эмбриологии.
ОГЛАВЛЕНИЕ
ОСНОВЫ ОБЩЕЙ ЭМБРИОЛОГИИ ……………………………………………4
Прогенез
Оплодотворение
Дробление
Бластуляция Гаструляция
ОСНОВЫ СРАВНИТЕЛЬНОЙ ЭМБРИОЛОГИИ……………………………….11
Особенности развития ланцетника
Особенности развития амфибий
Особенности развития рыб
Особенности развития птиц
Дифференцировка зародышевых листков
Эмбриональное развитие плацентарных млекопитающих и человека
Внезародышевые органы у плацентарных млекопитающих и человека
ЧАСТНАЯ ГИСТОЛОГИЯ
ЭМБРИОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ И ОРГАНОВ ЧУВСТВ……….….21
Развитие спинного мозга
Развитие спинномозговых и вегетативных узлов
Развитие головного мозга
Развитие гипофиза
Развитие органа зрения
Развитие органа слуха
ЭМБРИОГЕНЕЗ СЕРДЕЧНО - СОСУДИСТОЙ СИСТЕМЫ………………...29
Развитие сердца
Развитие артерий
Развитие вен
Особенности кровообращения плода
ЭМБРИОГЕНЕЗ ПИЩЕВАРИТЕЛЬНОЙ И ДЫХАТЕЛЬНОЙ СИСТЕМ…..32
Развитие ротовой полости и образование лица
Жаберный аппарат
Производные жаберного аппарата
Развитие кишечной трубки
Развитие печени и поджелудочной железы
Бранхиогенная группа желез
Развитие дыхательной системы
ЭМБРИОГЕНЕЗ ОРГАНОВ МОЧЕВЫДЕЛИТЕЛЬНОЙ СИСТЕМЫ……….37
ЭМБРИОГЕНЕЗ ПОЛОВОЙ СИСТЕМЫ ………………………………………...42
Индифферентная стадия
Развитие мужской половой системы
Развитие женской половой системы
ЛИТЕРАТУРА…………………………………………………………………….45
ОСНОВЫ ОБЩЕЙ ЭМБРИОЛОГИИ
Эмбриология - наука о закономерностях развития животных и человека от оплодотворения до приобретения зародышем структурных черт взрослого организма и способности к самостоятельному питанию. Изучение основ эмбриологии человека, знание физиологической динамики развития, его особенностей в различные сроки эмбрионального периода позволяют врачу понять и по возможности скорректировать патологию развития, а в ряде случаев даже осуществить ее профилактику.
В процессе эмбрионального развития человека сохраняются общие закономерности развития и стадии, характерные для позвоночных животных. Вместе с тем существуют особенности, отличающие развитие человека от развития других представителей позвоночных.
Прежде чем перейти к характеристике эмбриогенеза отметим, что в развитии эмбриологии как науки значительную роль сыграли отечественные исследователи. Так, в начале XIX века появились груды академиков Петербургской медицинской академии Каспара Фридриха Вольфа и Карла Максимовича Бэра. К. Ф. Вольф детально описал развитие птиц на примере цыпленка, а К. М. Бэр впервые описал строение яйцеклетки у человека. В середине XIX века работами Ильи Ильича Мечникова по развитию беспозвоночных и Александра Онуфриевича Ковалевского по эмбриогенезу ланцетника была доказана общность развития животных, что явилось основой для обоснования биогенетического закона о единстве их развития. Это позволяет считать И.И. Мечникова и А. О. Ковалевского основоположниками сравнительной эмбриологии.
Эмбриональное развитие человека принято делить на три периода: начальный (1-я неделя), зародышевый (2 – 8 неделя), плодный (с 9-й недели до рождения ребенка).
Эмбриогенез человека – часть его онтогенеза (индивидуальное развитие организма), включающая следующие основные стадии: 1 – оплодотворение и образование зиготы; II – дробление и образование бластулы (бластоцисты); III – гаструляцию (образование зародышевых листков); IV – гистогенез и органогенез зародышевых и внезародышевых органов; V - системогенез.
Сравнительная эмбриология, один из разделов эмбриологии, в сравнительном плане рассматривает развитие зародышей животных, стоящих на различных ступенях эволюции. Эти данные позволяют выделить основные этапы эмбриогенеза, общие для всех животных и человека, лучше определить место человека в эволюционном ряду, это особенно важно, если учесть, что невозможность экспериментирования на людях ограничивает возможности получения материала, касающегося эмбриогенеза человека. Помимо этого, ряд ранних стадий развития зародыша человека проходят очень быстро и не в полном объеме. Все это обуславливает необходимость для лучшего понимания эмбриогенеза человека, рассмотреть в сравнительном аспекте развитие более простых животных.
ПРОГЕНЕЗ
Эмбриогенез тесно связан с прогенезом (развитие и созревание половых клеток) и ранним постэмбриональным периодом.
С п е р м а т о з о и д - конечная клетка сперматогенеза – является во многих отношениях уникальной. Она резко вытянута и высокоспециализирована по всей своей длине. Головка сперматозоида содержит ядро, около двух третей которого покрыто акросомой. Основной компонент головки - клеточное ядро с гаплоидным набором конденсированных хромосом (23 хромосомы, одна из которых является половой). Плотность упаковки хромосом так велика, что объём ядра оказывается в 30 раз меньше, чем у гаплоидных клеток, образующихся сразу после мейоза. Это достигается за счёт того, что основными ядерными белками являются не гистоны, а другие основные белки, богатые аргинином и цистеином. В таком конденсированном состоянии генетический материал более защищён от повреждений при прохождении сперматозоида через оболочки яйцеклетки.
Спереди большая часть ядра, как двойной шапочкой, покрыта акросомой - это уплощённый пузырёк, происходящий из комплекса Гольджи и во многом сходный с лизосомой. Он содержит ферменты, необходимые для проникновения сперматозоида в яйцеклетку (коллагеназу, гиалуронидазу, акрозин, кислую фосфатазу и др.). Вся головка сперматозоида в целом, как и его хвост, окружена плазматической мембраной. В области головки эта мембрана содержит специальные белки. Одни из них заряжены отрицательно и способствуют (на малых расстояниях) направленному движению сперматозоида к яйцеклетке. Другие белки участвуют в связывании с яйцеклеткой. Постакросомальная зона головки сперматозоида является тем участком, цитоплазматическая мембрана которого при оплодотворении сливается с мембраной яйцеклетки.
Хвостовая часть сперматозоида наиболее протяженная и состоит из шейки, промежуточной, главной и терминальной (концевой) частей. Короткая шейка содержит 2 центриоли. Проксимальная центриоль прилежит к ядру, а от дистальной начинается осевая нить или аксонема, продолжающаяся во всех остальных частях хвостового отдела клетки.. Аксонема имеет обычное для всех жгутиков и ресничек строение, т.е. образована микротрубочками по схеме (9 х 2) + 2. Микротрубочки взаимодействуют с помощью динеиновых ручек; за счёт этого происходит скольжение соседних дуплетов друг относительно друга, приводящее к биению хвоста.
Вокруг аксонемы в промежуточной части хвоста находятся 9 пар периферических микротрубочек (играющих роль пассивных эластических структур) и “митохондриальное влагалище”, образованное митохондриями, расположенными по спирали, витки которой плотно прилегают друг к другу.
В главной части хвоста вокруг аксонемы рапсположены 9 пар наружных фибрилл и волокнистая оболочка (тонкофибриллярное влагалище).
Наконец, в концевой части хвоста аксонема покрыта непосредственно плазматической мембраной (имеющейся и в остальных отделах хвоста) и содержит единичные сократительные филаменты.
Я й ц е к л е т к а имеет шаровидную форму, больший, чем у сперматозоида, объем цитоплазмы, не обладает способностью самостоятельно передвигаться.
Классификация яйцеклеток основывается на признаках наличия, количества и распределения питательных веществ, расходующихся лишь после оплодотворения. Главные из них – желток, жир и гликоген. Желток представляет собой кристаллизованные вещества белковой природы, обычно в форме фосфопротеинов. Кристаллы выглядят как гранулы или пластинки различной формы, окруженные мембраной. Количество желтка, отлагающегося в яйцеклетке, сильно варьирует в разных группах животных. Количество желтка строго детерминировано генетически и не зависит от условий питания женской особи. Кроме того, желток содержит витамины А, В1, В2, , Д и Е. Большую часть жировых веществ составляют нейтральные жиры, а остальную – фосфатиды, фосфолипиды и холестерин.
Различают полилецитальные яйцеклетки, или многожелтковые (большинство членистоногих, рыбы, птицы), мезолецитальные - со средним количеством желтка (амфибии, осетровые рыбы), олиголецитальные - маложелтковые (беспозвоночные. млекопитающие) и алецительные - безжелтковые (плоские черви). Маложелтковые яйцеклетки подразделяются на первичные (ланцетник) и вторичные (у плацентарных млекопитающих и человека). Как правило, в маложелтковых яйцеклетках желточные включения распределены более или менее равномерно. Такая яйцеклетка носит название изолецитальной. У плацентарных млекопитающих и у человека, питание зародыша идет за счет материнского организма, поэтому нет необходимости запасать питательный материал в яйцеклетке, но для отличия от аналогичной по структуре яйцеклетки низших хордовых, яйцеклетка млекопитающих носит название вторично изолецитальной.
Для развития более высокоорганизованных животных, исключая млекопитающих, требуется больше питательных веществ. Полилецитальные яйцеклетки могут быть по распределению желтка центролецитальными (членистоногие) и телолецитальными (рыбы, кроме осетровых, амфибии). В процессе отложения желтка выявляется полярность яйцеклетки, которое проявляется накоплением питательных веществ на одном из полюсов (вегетативный полюс), в то время, как противоположный (анимальный полюс) содержит зародышевый материал. Такие яйцеклетки получили название телолецитальных. При этом у одних животных телолецитальные клетки имеют слабо выраженную полярность (амфибии), у других животных телолецитальные клетки характеризуются резко выраженной полярностью (рыбы, птицы).
Яйцеклетка человека достигает в диаметре 130 мкм. Вокруг её цитолеммы располагается блестящая, или прозрачная, зона (zona pellucida), состоящая из гликопротеинов и гликозаминогликанов. Блестящую зону окружает слой фолликулярных клеток, выполняющих трофическую и защитную функции. В цитоплазме развиты органеллы синтеза белка и аппарат Гольджи, продуцирующие кортикальные гранулы и занимающие периферическую часть цитоплазмы. Кортикальные гранулы содержат гликозаминогликаны и протеолитические ферменты, участвуют в кортикальной реакции.
ОПЛОДОТВОРЕНИЕ
Развитие зародыша начинается с процесса оплодотворения, в ходе которого мужская и женская клетки, сливаясь, образуют зиготу, которая качественно отличается от половых клеток тем, что содержит диплоидный набор хромосом.
Оплодотворением называют слияние сперматозоида с яйцеклеткой, завершающееся объединением их ядер в единое ядро оплодотворенного яйца – зиготы. Сперматозоид выполняет две функции. Первая – активация яйцеклетки, побуждение его к началу развития. Развитие яйцеклетки без участия сперматозоида называется партеногенезом. Другая функция сперматозоида – внесение в яйцеклетку генетического материала.
Взаимодействие половых клеток можно разделить на три фазы: 1) дистантное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 3) вхождение сперматозоида в яйцеклетку и последующее слияние – сингамия.
Дистантное взаимодействие гамет. После эякуляции сперматозоиды под действием секрета женских половых путей лишаются гликопротеинов и протеинов семенной плазмы в области акросомы. Такое явление называется капацитацией - приобретение спермиями оплодотворяющей способности. Основная биологическая задача гамет – обеспечить встречу половых клеток и вместе с тем, у ряда видов, предохранить яйцеклетку от проникновения в нее излишнего количества сперматозоидов. У некоторых видов на этом этапе распространено явление хемотаксиса – движение сперматозоидов вверх по градиенту концентрации некоторых веществ, выделяемых яйцеклеткой (костистые рыбы). В большинстве же случаев дистантные влияния яйцеклетки на сперматозоид не носят направленного характера. Они осуществляются двумя классами веществ, называемых гиногамоны I и II. Гиногамоны I - низкомолекулярные вещества небелковой природы, выделяемые яйцевыми оболочками. Эти вещества активируют движения спермиев. Гиногамоны II (фертилизины, изоаглютинины) - гликопротеиды, вызывающие склеивание сперматозоидов путем иммунной реакции с андрогамоном II (антифертилизином), молекулы которого встроены в оболочку сперматозоида. В свою очередь, сперматозоиды выделяют во внешнюю среду андрогамон I – вещество небелковой природы, являющееся антагонистом гиногамона I и подавляющее подвижность спермиев.
Контактное взаимодействие гамет – вторая фаза оплодотворения, во время которой сперматозоиды приближаются к яйцеклетке и вступают в контакт с ее оболочкой. Яйцеклетка начинает вращаться вокруг своей оси, что обусловлено биением жгутиков спематозоидов. В процессе взаимодействия мужской и женской половых клеток в сперматозоидах происходит акросомальная реакция, которая характеризуется повышением проницаемости плазмолеммы сперматозоида к ионам Са2+ и ее деполяризацией. Это способствует слиянию плазмолеммы с передней мембраной акросомы и вещества акросомы (спермолизины) выходят в окружающую среду, что приводит к быстрому разрыхлению яйцевых оболочек в месте контакта со спермием. Ферменты разрушают блестящую зону, спермий проходит через нее и входит в перивителлиновое пространство, расположенное между блестящей зоной и плазмолеммой яйцеклетки. Через несколько секунд изменяются свойства плазмолеммы яйцеклетки и начинается кортикальная реакция. Кортикальная реакция – слияние плазмолеммы яйцеклетки с мембраной кортикальных гранул, в результате чего содержимое из гранул выходит в перивителлиновое пространство и воздействуют на молекулы гликопротеидов блестящей зоны. В это же время возникают выпячивания мембраны акросомы, в результате чего образуются акросомные трубочки. Они удлиняются и контактируют с плазматической мембраной яйцеклетки. Мембраны клеток сливаются, ядро и центриоли сперматозоида по акросомным трубочкам переходят в яйцеклетку.
Вхождение сперматозоида в яйцеклетку. В ооплазму проникают головка и промежуточная часть хвостового отдела. Кортикальная реакция заканчивается выделением фактора, способствующим затвердению прозрачной зоны и образованию из нее оболочки оплодотворения, препятствующей полиспермии – проникновению других сперматозоидов. Проникший в яйцеклетку сперматозоид сразу же поворачиваеися шейкой вперед. Хроматин в ядре сперматозоида деспирализуется. Ядро сперматозоида теперь называется мужским пронуклеусом. Аналогичные процессы проходят в ядре яйцеклетки. Это ядро называется женским пронуклеусом. Центриоли сперматозоида становятся центром движения внутри оплодотворенной яйцеклетки (зиготы). Объединение двух пронуклеусов – синкарион.
В процессе эмбрионального развития из оплодотворенной яйцеклетки возникает многоклеточный организм с различными органами и тканями. Следовательно, в основе эмбрионального развития должны лежать процессы: 1) размножения клеток, 2) специализации клеток и образуемых ими тканей в разных направлениях (дифференцировка), 3) роста и 4) перемещения отдельных клеток и клеточных масс, что приводит к формированию зачатков органов.
ДРОБЛЕНИЕ
Образовавшаяся зигота начинает постепенно превращаться в многоклеточный организм путем митотического деления. Однако, дочерние клетки, их называют бластомерами, не успевают вырасти до размеров материнских, что приводит к постоянному уменьшению в размерах образующихся новых бластомеров, поэтому данный этап эмбриогенеза зародыша получил название - дробление. Клетки, образующиеся в результате нескольких первых делений дробления, по большей части неспециализированы. Их синтетическая активность направлена на вырабатывание ДНК и белков, необходимых для процессов клеточного деления, а не для специализированной бмохимической активности. Процесс дробления протекает по-разному у разных животных. Один из главных факторов, определяющих эти различия – количество содержащегося в яйце желтка.
Указанные особенности строения яйцеклетки определяют характер дробления. Деление олиголецитального яйца по существу ничем не отличается от обычного митоза, в дроблении участвует вся зигота. Образующиеся дочерние клетки имеют одинаковые размеры, но они мельче материнской клетки. Такой тип дробления называется полным (голобластическим). Так делятся изолецитальные и телолецитальные яйцеклетки с нерезко выраженной полярностью. Если образующиеся бластомеры одинаковы по размеру, то говорят о равномерном дроблении (ланцетник), если же бластомеры различаются по величине - дробление неравномерное (амфибии, млекопитающие). Полилецитальные яйцеклетки обуславливают неполное (меробластическое, дискоидальное) дробление зиготы. При этом дробится лишь материал анимального полюса. После слияния пронуклеусов ядро зиготы делится на много ядер, которые по цитоплазматическим мостикам перемещаются (“выползают”) во внешний слой свободной от желтка цитоплазмы, образуя там клеточный слой – бластодерму. В полилецитальных яйцеклетках как центро-, так и телолецитального типа свободная от желтка цитоплазма расположена тонким слоем. Огромный по сравнению с чистой цитоплазмой объем желтка (материал вегетативного полюса) не делится. Такой тип дробления называется частичным, неполным или меробластическим. Для ряда полилецитальных яйцеклеток харктерен дискоидальный тип дробления (рыбы, птицы) – деление на бластомеры лишь тонкого диска цитоплазмы, расположенного на анимальном полюсе. Дробление зиготы разных видов отличается и скоростью деления бластомеров, что позволяет выделять синхронный и асинхронный типы дробления. Синхронное или правильное дробление характеризуется одинаковой скоростью во всех участках зародыша, что обеспечивает увеличение количества бластомеров в правильной геометрической прогрессии (2,4,8,16 и т.д.). Асинхронное или неправильное дробление характеризуется разной скоростью дробления в различных участках зародыша, и правильность увеличения количества бластомеров нарушается.
После нескольких клеточных делений, каждое из которых приводит к удвоению числа клеток в зародыше, зародыш выглядит как небольшая тутовая ягода – морула (лат. morula - тута).
БЛАСТУЛЯЦИЯ. ТИПЫ БЛАСТУЛ.
У большинства яиц еще на ранних стадиях дробления внутренние стенки бластомеров начинают расходиться, расслаиваться и между ними возникает сначала небольшая, а затем все увеличивающаяся полость дробления (бластоцель). Бластомеры перестают быть округлыми и увеличивают поверхности взаимного контакта. В связи с этим стенка зародыша эпителизуется – приобретает вид эпителиального пласта. Эпителизация – одна из важнейших предпосылок более тесных взаимодействий между клетками и их скоординированных движений в ходе дальнейшего развития. На этом этапе развития зародыш называется бластулой, а процессы формирующие бластулу – бластуляцией. Бластула имеет стенку – бластодерму и полость – бластоцель., заполненную жидкостью – продуктом секреции бластомеров. В бластодерме различают крышу (материал анимального полюса), дно (материал вегетативного полюса) и краевую зону, расположенную между ними.
Строение бластулы зависит от типа дробления данного яйца. В олиголецитальных яйцеклетках, при полном равномерном дроблении (ланцетник, иглокожие), бластула имеет тонкую однослойную стенку, а обширная бластоцель находится в центре. Такая бластула называется целобластулой.
При полном неравномерном дроблении (миноги, лягушки) образуется амфибластула -бластула с многослойной бластодермой и эксцентрично расположенным бластоцелем. Крыша такой бластулы состоит из мелких бластомеров, а дно – из крупных, богатых желтком, бластомеров.
В полилецитальных яйцах рыб и птиц дробление дискоидальное. В результате образуется состоящий из нескольких клеточных слоев диск, лежащий на нераздробленной массе желтка. Диск несколько выгибается над желтком, и между ними возникает полость – бластоцель. Это дискобластула.
При полном и асинхронном дроблении (млекопитающие) образуется зародышевый пузырек – бластоциста. В нем различают уплощенный наружный слой клеток - т рофобласт, и скопление бластомеров в виде узелка на внутренней поверхности трофобласта – эмбриобласт. Переход от морулы к бластуле у зародышей млекопитающих происходит очень быстро. В центре зародыша образуется большая, наполненная жидкостью, полость – бластоцель. Эта жидкость секретируется бластомерами. Уплощаясь, клетки трофобласта образуют плотные межклеточные контакты, что сильно снижает проницаемость этого слоя, предотвращая свободное прохождение жидкости. На ранних стадиях бластоциста млекопитающих остается заключенной в zona pellucida (блестящая зона).
ГАСТРУЛЯЦИЯ
Общая сущность этого процесса у всех многоклеточных животных состоит в том, что однослойный зародыш – бластула, представленный только одним клеточным пластом – бластодермой, превращается в двуслойный, а у позвоночных затем и в трехслойный зародыш – гаструлу, состоящую из наружного зародышевого листка – эктодермы и внутреннего зародышевого листка – энтодермы. У позвоночных уже в ходе гаструляции возникает третий, средний зародышевый листок – мезодерма. Появление зародышевых листков – первый этап дифференциации клеточного материала зародыша. Далее из этих листков развиваются осевые зачатки органов. Образование некоторых осевых зачатков (хорды) идет почти одновременно с гаструляцией. Гаструляция происходит под влиянием организующих факторов – индукторов, влияющих на участки зародыша и вызывающие их развитие в определенном направлении. Индуцирующими факторами могут быть белки, нуклеопротеиды, стероиды и другие химические вещества.
Гаструляция может совершаться различными способами. Способ гаструляции во многом определяется типом бластулы, который, в свою очередь, обусловлен типом исходной яйцеклетки.
1. Инвагинация или впячивание бластодермы вегетативного полюса по направлению к анимальному. Зародыш принимает вид чаши с двухслойной стенкой. Внутренний слой формирует первичную энтодерму, а наружный – первичную эктодерму. Полость гаструлы, появившаяся в результате впячивания энтодермы, называется гастроцель. Отверстие, которым гастроцель сообщается с внешней средой, называется первичным ротовым отверстием – бластопором. Такой способ гаструляции распространен у иглокожих и низших хордовых (ланцетник).
2. Эпиболия - нарастание анимальной части бластулы на вегетативную и, таким образом, переход вегетативной части во внутреннее положение. Материал, оставшийся на поверхности зародыша после завершения гаструляции, является эктодермой, а погрузившийся внутрь - энтодермой. Эти два способа гаструляции часто сопровождают друг друга. В результате эпиболии к краям бластопора подводятся клетки, которым предстоит инвагинировать, вворачиваясь через губу бластопора, чтобы занять свое место в одной из внутренних структур зародыша. У некоторых животных гаструляция может осуществляться комбинацией двух или более различных способов, например инвагинации и эпиболии у амфибий.
3. Деламинация – расслоение, расщепление единого пласта на два – наружный (зктодерму) и внутренний (энтодерму). Такой способ обособления энтодермы наблюдается у многих беспозвоночных (членистоногие) и у высших позвоночных (птицы, плацентарные млекопитающие, человек.)
4. Иммиграция – состоит в том, что часть клеток бластодермы выселяется, уходит в бластоцель и там складывается во второй, внутренний зародышевый листок – энтодерму. Клетки, оставшиеся на месте, образуют эктодерму. Такой тип гаструляции был открыт И.И. Мечниковым в 1886 году и считается эволюционно наиболее древним. Этот способ распространен у кишечнополостных. Выселение клеток характерно и для высших позвоночных (птицы), плацентарных млекопитающих и человека. В этом случае она составляет второй этап гаструляции и приводит к образованию среднего зародышевого листка – мезодермы.
Развитие всех низших позвоночных происходит в воде. Все их представители откладывают яйца в воду, где и происходит эмбриональное и личиночное развитие.
Высшие позвоночные полностью перешли к сухопутному образу жизни и стали откладывать яйца на суше. Это обстоятельство наложило свой отпечаток на все эмбриональные процессы у этих животных. Повышение уровня организации животного потребовало увеличение количества желтка, которым снабжается яйцо во время своего формирования в яичнике. Это в свою очередь повлекло изменения дробления и способа гаструляции. Развитие в воздушной, а не в водной среде, потребовало развития специальных приспособлений, защищающих зародыш от высыхания, механических повреждений и др. (яйцевые и зародышевые оболочки). Появление толстых и прочных оболочек (скорлуповая, серозная оболочки) на яйцевых клетках рептилий и птиц, сделало невозможным наружное оплодотворение. Возникла необходимость проникновения сперматозоидов в яйцеклетку на такой стадии развития, когда этих оболочек еще нет. Следовательно, единственно возможным сделалось внутреннее осеменение и оплодотворение.
ОСНОВЫ СРАВНИТЕЛЬНОЙ ЭМБРИОЛОГИИ
ОСОБЕННОСТИ РАЗВИТИЯ ЛАНЦЕТНИКА
Яйцеклетка ланцетника первично изолецитальная. Дробление зиготы голобластическое, равномерное, синхронное.
Для ланцетника характерно образование целобластулы. Снаружи в бластуле выделяют крышу (верхняя часть), дно (нижняя часть) и краевую зону (лежит на границе между крышей и дном).
Гаструляция путем инвагинации (впячивание дна бластомера) и образования двуслойной чашеобразной гаструлы, чья гастроцель сообщается с внешней средой широким бластопором, соответствующим будущему каудальному концу зародыша. Впятившиеся клетки дна образуют внутренний зародышевый листок – энтодерму, тогда как наружная стенка гаструлы оказывается образованной клетками крыши, или наружным зародышевым листком – эктодермой. Образование экто- и энтодермы знаменует собой процесс ранней гаструляции. Образование третьего листка (мезодермы) происходит путем выпячивания части энтодермы в виде двух карманов, внедряющихся между двумя имеющимися листками. В дальнейшем,уже за счет имеющихся зародышевых листков, формируются осевые органы: хорда, нервная и кишечная трубки.
В дальнейшем мезодермальные пузыри (крылья) делятся на дорзальную часть (миотом) и вентральную (спланхнотом). Дорсальные отделы мезодермы вдоль длинника всего тела сегментируются на сомиты (10 – 11 пар, у человека – 44 пары сомитов), за исключением хвостовой части. Спланхнотом не сегментируется и имеет в своем составе два листка, ограничивающих вторичную полость тела – целом: париетальный (прилежит к эктодерме) и висцеральный (прилежит к энтодерме). На границе между дорсальной и вентральной частями находится скопление клеток, называемое нефротомом, который, как и миотом, сегментирован в головной и туловищной частях (сегментные ножки), но не имеет сегментации в хвостовом отделе (нефрогенные тяжи).
Зародышевые листки и осевые органы соединены между собой за счет подвижных отростчатых клеток, выселяющихся из разных отделов мезодермы (главным образом из сомитов и спланхнотома) и формирующих мезенхиму. После своего возникновения мезенхима распространяется между всеми органами зародыша и образует ту внутреннюю среду, через которую совершается обмен веществ между отдельными органами.
За счет образовавшихся зародышевых листков (экто-, энто-, мезодермы), осевых органов (нервной и кишечной трубки) и мезенхимы формируются все системы органов, имеющихся в теле животного.
ОСОБЕННОСТИ РАЗВИТИЯ АМФИБИЙ
Яйцеклетка – телолецитальная с нерезко выраженной полярностью. Дробление голобластическое, неравномерное, асинхронное. Морула содержит крупные бластомеры. Бластула имеет многослойную бластодерму, бластоцель смещена к анимальному полюсу, где сконцентрированы более мелкие бластомеры – амфибластула. Гаструляция начинается путем инвагинации, что приводит к образованию удлиненного образования – углубления (серповидная бороздка) на границе анимального и вегетативного полюсов. В дальнейшем гаструляция у амфибий продолжается путем обрастания бластомерами анимального полюса вегетативной части зародыша. Такой вариант гаструляции получил название эпиболии. В итоге, вегетативная часть оказывается внутри зародыша, заполняя около половины внутреннего пространства сферической гаструлы. А в стенке последней содержатся все три зародышевых листка. Гаструляция амфибий характеризуется и тем, что все зародышевые листки образуются практически одновременно и быстро дифференцируются.
ОСОБЕННОСТИ РАЗВИТИЯ РЫБ
Яйцеклетка – телолецитальная с резко выраженной полярностью. Дробление меробластическое, неравномерное, асинхронное. В результате дробления образуется не шаровидная морула, а зародышевый диск уплощенной формы. Затем зародышевый диск преобразуется в дискобластулу. Первая фаза гаструляции у разных представителей рыб происходит либо путем деламинации, либо путем инвагинации. Вторая фаза гаструляции – иммиграция.
Характерной особенностью развития рыб является, в отличие от ланцетника и амфибий, формирование внезародышевого органа – желточного мешка. Его формирование начинается на стадии ранней гаструлы, когда во внутреннем листке можно выделить зародышевую (кишечную) энтодерму и расположенную по периферии диска внезародышевую (желточную) энтодерму. Развивается желточный мешок за счет постепенного обрастания желтка краем зародышевого диска (внезародышевые части энтодермы и висцерального листка мезодермы обрастают желток, формируя желточный мешок). На периферии зародыша к этим листкам присоединяются париетальный листок мезодермы и эктодерма. Поступление питания к зародышу происходит за счет образующихся в стенке желточного мешка кровеносных сосудов, подходящих к зародышу через желточный стебелек, связывающий зародыш с желточным мешком. Желточный мешок выполняет у рыб трофическую и кроветворную функции. У высших представителей позвоночных, в связи с изменениями условий развития, меняются и его функции.
ОСОБЕННОСТИ РАЗВИТИЯ ПТИЦ
Яйцеклетка телолецитальная с резко выраженной, полярностью. Особенностями ее строения являются наличие ряда оболочек (желточная, белковая, две подскорлуповых, между которыми на одном из полюсов образуется воздушная камера, и скорлупа), отражающих то, что развитие зародыша идет не в водной, а в воздушной среде.
Дробление у птиц меробластическое, неравномерное, асинхронное. Результат дробления – зародышевый диск, а затем – дискобластула. Деламинация происходит на первом этапе гаструляции. При этом одноклеточный слой дискобластулы расщепляется путем деления клеток на два слоя: верхний – эпибласт и нижний – гипобласт. Гипобласт образован плоскими клетками, покрывает желток и в формировании тканей зародыша не участвует. Эпибласт состоит из цилиндрических клеток и из его центральной части (зародышевый щиток) формируются все три зародышевых листка – эктодерма, мезодерма и энтодерма. Иммиграция клеток зародышевого эпибласта и формирование мезодермы и осевых органов происходит так же, как у млекопитающих и человека.
Для эмбриогенеза птиц характерно развитие не только желточного мешка, но еще ряда других внезародышевых органов: амниона, серозы, аллантоиса.
Амнион (водная оболочка) и серозная оболочка образуются за счет выпячивающейся над зародышем эктодермы и париетального листка мезодермы (спланхнотома), формируются амниотические складки. Указанные складки наползают (сверху и сзади) на зародыш и смыкаются над ним, образуя амниотическую оболочку, которая заполняется в дальнейшем жидкостью. Эта жидкость вырабатывается клетками эктодермы амниотической оболочки и содержит белки, углеводы. В то же время наружные части амниотической складки обрастают весь зародыш и желточный мешок, то есть, происходит обрастание теми же листками внутренней поверхности скорлупы яйца, формируя еще один внезародышевый орган – серозную оболочку, которая участвует в газообмене, снабжая эмбрион кислородом (провизорный орган дыхания). Амнион выполняет трофическую и защитную функции, окружая зародыш жидкой средой, обеспечивает условия для свободного развития зародыша и предохраняет его от вредных внешних воздействий.
Аллантоис развивается как выпячивание из вентральной стенки задней кишки, образованной энтодермой и висцеральным листком мезодермы. Аллантоис имеет удлиненную форму и, разрастаясь, внедряется в щель между амнионом и серозной оболочкой, подрастая к воздушной камере яйца. Аллантоис у птиц участвует в обмене газов и является резервуаром, куда в процессе эмбриогенеза попадают продукты, подлежащие выделению. По сосудам, образующимся в мезодерме аллантоиса, доставляется кислород. В аллантоис выделяются продукты обмена веществ зародыша.
ДИФФЕРЕНЦИРОВКА ЗАРОДЫШЕВЫХ ЛИСТКОВ.
Дифференцировка – составная часть гаструляции и проявляется в том, что клетки приобретают все большие биохимические и морфологические различия друг от друга, а возможности их дальнейшего развития сужаются. В то же время, дифференцировка происходит на всех этапах эмбриогенеза (начиная с зиготы) и продолжается также во взрослом организме. 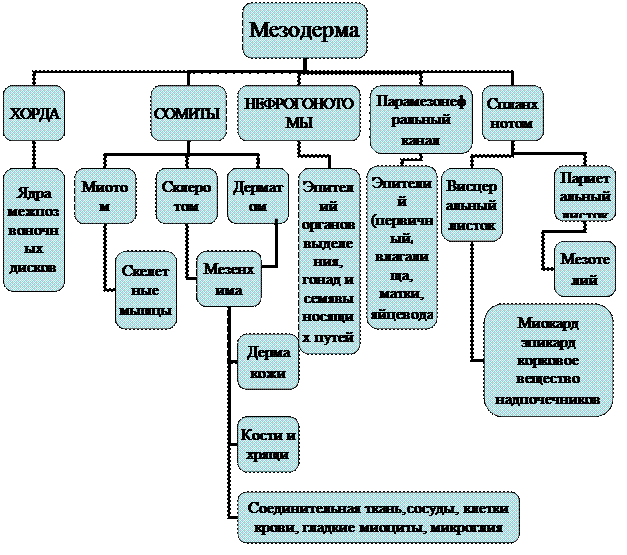

ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ПЛАЦЕНТАРНЫХ МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА
Эмбриогенез млекопитающих и человека резко отличается от эмбриогенеза других классов позвоночных. Это связано с тем, что развитие их зародыша происходит в организме матери.
Начальные этапы эмбрионального развития у млекопитающих и человека протекают одинаково.
Яйцеклетка вторично изолецитальная. Оплодотворение – внутреннее; происходит впроксимальной трети маточной трубы и, здесь же, начинается дробление зиготы. Дробление голобластическое, неравномерное и асинхронное. Первое же дробление дает два бластомера неодинаковой величины. Один из бластомеров светлый и мелкий, второй – темный и крупный. В течение первых двух – трех суток дробление протекает медленно, зародыш продвигается по маточным трубам вследствие тока жидкости, создаваемым сокращениями мускулатуры яйцевода и мерцанием ресничек его эпителия. Начиная с трех суток, дробление идет быстрее, и на 4-е сутки зародыш состоит из 7 – 12 бластомеров. При последующих дроблениях количество светлых бластомеров резко увеличивается и они окружают сплошным слоем темные бластомеры, которые оказываются расположенными внутри в виде плотного скопления. Слой светлых бластомеров, окружающий темные, носит название трофобласта. Он выполняет трофическую функцию, всасывая серозную жидкость из просвета яйцевода, а позже – из полости матки и обеспечивает питательными веществами скопление темных бластомеров, называемое эмбриобластом. На описываемом этапе развития зародыш нередко называют стереобластулой или морулой, а на 3 – 4-е сутки начинается формирование бластоцисты – полого пузырька, заполненного жидкостью. Вследствие накопления под слоем трофобласта жидкости, эмбриобласт оттесняется, располагается у одного полюса в виде зародышевого узелка, который изнутри прикрепляется к трофобласту и начинается процесс имплантации. Зародышевый узелок уплощается и превращается в зародышевый щиток, в котором начинается подготовка к первой стадии гаструляции.
До стадии бластоцисты развитие плацентарных млекопитающих и человека идет почти одинаково. В дальнейшем ходе развития, появляется ряд отличий.
Ипмплантация – внедрение зародыша в слизистую оболочку стенки матки. В процессе имплантации различают две стадии: 1) адгезию или прилипание (зародыш прикрепляется к внутренней поверхности матки (эндометрию), и 2) инвазию или погружение (зародыш внедряется в ткани слизистой оболочки матки). После адгезии клетки трофобласта начинают быстро пролиферировать (делиться).
На 5 – 6 день бластоциста оказывается в полости матки в свободном виде, и эта стадия обозначается как свободная бластоциста, окруженная оболочкой оплодотворения.
Полость бластоцисты человека заполнена серозной жидкостью. Из эмбриобласта выделяются клетки внезародышевой мезодермы (мезенхимы) которые принимают отростчатую форму и выстилают изнутри трофобласт, а также окружают со всех сторон эмбриобласт. К 11-м суткам внезародышевая мезодерма заполняет полость бластоцисты. На 7-е сутки эмбрионального развития начинается процесс имплантации (внедрение зародыша в ткань слизистой оболочки матки). В трофобласте и эмбриобласте происходят изменения, связанные с подготовкой к этому процессу. Основная роль в этом процессе принадлежит трофобласту, в котором увеличивается количество лизосом с ферментами, обеспечивающее разрушение тканей матки (эпителий и соединительная ткань) и тем самым, способствующим внедрению зародыша в толщу ее слизистой оболочки. Погружающийся зародыш образует в эндометрии имплантационную ямку и полностью оказывается в толще эндометрия. К этому времени трофобласт за счет размножения клеток становится двухслойным. Внутренний слой представлен кубическими клетками и носит название цитотрофобласт, а наружный слой утрачивает клеточное строение и представляет собой общую массу цитоплазмы с находящимися в ней делящимися ядрами – хориальный синцитий или симпластотрофобласт, продуцирующий протеолитические ферменты. Появляющиеся в трофобласте микроворсинки (будущие ворсинки хориона) постепенно разрушают оболочку оплодотворения. После полного погружения зародыша дефект в эндометрии вначале заполняется свернувшейся кровью. Затем зарастает соединительной тканью и к 12 – 13 дню покрывается эпителием. Вокруг зародыша образуются лакуны, заполняющиеся материнской кровью. В клетках соединительной ткани развивается децидуальная реакция: разрастаются сосуды, появляется отечность, клетки увеличиваются в объеме и накапливают гликоген и липиды.
Первые 2 недели трофобласт потребляет продукты распада материнских тканей - это гистиотрофный тип питания. Затем питание зародыша осуществляется непосредственно из материнской крови – гематотрофный тип питания. Питание и дыхание зародыша происходит за счет питательных веществ и кислорода, находящихся в крови матери. Продукты обмена веществ зародыша выделяются также в кровь материнского организма. Далее идет формирование плаценты гемохориального типа, обеспечивающей наиболее совершенный уровень обмена веществ зародыша.
Параллельно с имплантацией зародыша и формированием хориона, в материале зародышевого щитка начинаются процессы гаструляции. Гаструляция у человека осуществляется в две фазы.
Первая фаза гаструляции у человека происходит на 7-е сутки по типу деляминации, одновременно с имплантацией. Группа клеток эмбриобласта уплощается (напоминая зародышевый диск птиц) и эмбриобласт расщепляется на гипобласт (нижний, или внутренний листок) и эпибласт (верхний, или наружный листок). Гипобласт представлен мелкими, кубической формы, светлыми клетками, а эпибласт состоит из цилиндрических клеток, приобретающих эпителиоподобный вид, ядра которых лежат на разных уровнях (псевдомногорядность).
Гипобласт принимает участие в формировании внезародышевого органа – желточного мешка. Для этого он разрастается по внутренней поверхности трофобласта и формирует внезародышевую энтодерму.
У высших животных и у человека между клетками эпибласта появляются мелкие полости, которые затем сливаются в единую амниотическую полость. В конечном счете (после перемещения клеток крыши этой полости), эпибласт расщепляется еще на два листка – зародышевый эпибласт, образующий дно амниотического пузыря, и амниотическую (внезародышевую) эктодерму - крышу пузырька. Амниотическая эктодерма принимает участие в образовании внезародышевого органа – амниона. У других животных амнион формируется иначе.
Центральную роль в образовании тканей зародыша играет зародышевый эпибласт, который во второй фазе гаструляции, служит источником формирования всех трех зародышевых листков.
Между первой и второй фазами гаструляции идет образование внезародышевых органов, необходимых для успешного развития зародыша.
Таким образом, первая фаза гаструляции приводит к обособлению источника зародышевых листков (зародышевого эпибласта) и является началом формирования внезародышевых органов.
Вторая фаза гаструляции происходит путем иммиграции клеток. Начинается на 14-е сутки и продолжается до 17-х суток развития, в результате чего образуются мезодерма и комплекс осевых органов.
В результате миграции клеток в области эпибласта появляются: 1) первичная полоска – утолщение эпибласта с первичной бороздкой посередине; 2) первичный (гензеновский) бугорок – возвышение в передней части первичной полоски, в центре которого находится первичная ямка. Это места активной миграции клеток внутрь с внешней поверхности зародыша. Кроме того, в переднем участке эпибласта формируется еще одно утолщение – прехордальная пластинка. Затем клетки эпибласта начинают мигрировать через первичную полоску внутрь. Одни клетки, проходя через первичную полоску, оттесняют в стороны клетки гипобласта и дают начало зародышевой энтодерме. Другие клетки распространяются в виде рыхлого пласта между эпибластом и гипобластом (точнее, уже энтодермой) и дают начало зачатку хорды и зародышевой мезодерме, образуя мезодермальные крылья по бокам от хорды. Остающиеся на поверхности клетки эпибласта формируют зародышевую эктодерму. Гипобласт сохраняется и образует внезародышевую энтодерму, принимающую участие в формировании желточного мешка.
К 15-17 суткам зародыш имеет следующее строение. Трофобласт вместе с внезародышевой мезодермой образует хорион. В зародышевой части располагаются прилегающие друг к другу амниотический и желточный пузырьки. Эта часть зародыша прикреплена к хориону при помощи амниотической ножки, представляющей собой плотный тяж клеток внезародышевой мезодермы. Амниотическая ножка – зачаток пупочного канатика.
С 20-го дня внутриутробного развития мезодерма (с каждой стороны) дифференцируется на три части: дорзальную мезодерму, вентральную мезодерму (спланхнотом) и промежуточную мезодерму (нефротом). Далее, происходит выселение части клеток в основном из сомитов. Эти клетки приобретают звездчатую форму и заполняют промежутки между зародышевыми листками, образуя внезародышевую мезодерму или мезенхиму, называемую некоторыми авторами четвертым зародышевым листком.
Также, с 20 – 21 суток начинает обособляться тело зародыша. Зародышевый щиток как бы приподнимается над желточным мешком и сворачивается, отделяясь от него туловищной складкой. При этом зародышевая энтодерма замыкается в кишечную трубку, которая, однако, в среднем отделе еще имеет сообщение с желточным мешком. Кроме латерального сгибания, происходит еще и продольное сгибание зародыша, так что его передний и задний отделы сгибаются в сторону желточного мешка. В результате, прехордальная пластинка перемещается с дорсальной поверхности на вентральную. Именно здесь, при последующем разрыве этой пластинки, оказывается ротовое отверстие эмбриона.
В конечном счете, зародыш оказывается связанным с желточным мешком стебельком, который входит в состав амниотической ножки, а позднее – пупочного канатика. С этого момента амниотическая оболочка окружает весь зародыш (за исключением места отхождения амниотической ножки).
ВНЕЗАРОДЫШЕВЫЕ ОРГАНЫ У ПЛАЦЕНТАРНЫХ МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА
Внезародышевые органы, развивающиеся в процессе эмбриогенеза вне тела зародыша, выполняют многообразные функции, обеспечивающие рост и развитие самого зародыша. К ним относятся амнион, желточный мешок, аллантоис, хорион, плацента.
Амнион – временный орган, обеспечивающий водную среду для развития зародыша. Он возник в эволюции в связи с выходом позвоночных животных на сушу. Стенка амниотического пузырька изнутри покрыта внезародышевой (амниотической) эктодермой, снаружи - внезародышевой мезенхимы, которые формируют соединительную ткань органа. К концу 7-й недели развития зародыша, соединительная ткань амниона входит в контакт с соединительной тканью хориона. Эпителий амниона переходит на амниотическую ножку, превращающуюся в пупочный канатик.
Эпителий амниона на ранних стадиях – однослойный плоский, активно делящийся, который постепенно преобразуется в призматический. На поверхности эпителиальных клеток появляются микроворсинки. В цитоплазме содержатся небольшие капли липидов, зерна гликогена и гликозаминогликаны.
Основная функция амниона – выработка околоплодных вод, обеспечивающих среду для развивающегося организма и предохраняющих его от механического повреждения. Эпителий амниона принимает участие и в обратном всасывании околоплодных вод
Желточный мешок - возник как орган, депонирующий питательные вещества (желток), необходимые для развития зародыша. У человека он образован внезародышевой энтодермой и внезародышевой мезодермой (мезенхимой). В отличие от птиц, у человека и других млекопитающих желточный мешок практически не содержит желтка, а заполнен серозной жидкостью. Желточный мешок появляется на 2-й неделе развития и в питании зародыша участия практически не принимает, так как уже на 3-й неделе развития устанавливается связь плода с материнским организмом, и возникает гематотрофное питание. В стенке желточного мешка развиваются кровяные островки и первые кровеносные сосуды, обеспечивающие перенос кислорода и питательных веществ у плода. В качестве кроветворного органа желточный мешок функционирует до 7 – 8-й недели, а затем подвергается обратному развитию и остается в составе пупочного канатика в виде узкой трубочки, служащей проводником кровеносных сосудов к плаценте.
Аллантоис - это небольшой пальцевидный вырост в каудальном отделе кишечной энтодермы и висцерального листка мезодермы. Аллантоис является производным желточного мешка и состоит из внезародышевой энтодермы и висцерального листка мезодермы. Зародыш располагается между амниотическим и желточным пузырьками и связан с хорионом с помощью амниотческой ножки, в которую врастает аллантоис. Проксимальная часть аллантоиса располагается вдоль желточного стебелька, а дистальная, разрастаясь, врастает в щель между амнионом и хорионом. У человека аллантоис не достигает значительного развития, но обеспечивает питание и дыхание зародыша, так как по нему к хориону растут сосуды. Это орган газообмена и выделения. На
2-м месяце эмбриогенеза аллантоис редуцируется и превращается в тяж клеток, который вместе с редуцированным желточным мешком входит в состав пупочного канатика.
Пупочный канатик соединяет зародыш с плацентой. Он представляет собой упругий тяж, покрытый амниотической оболочкой. Основу пупочного канатика составляет слизистая волокнистая соединительная ткань - вартонов студень, образовавшийся из внезародышевой мезодермы амниотической ножки, где проходят две пупочные артерии и одна вена, а также рудименты желточного мешка и аллантоиса.
Слизистая соединительная ткань обеспечивает упругость канатика, предохраняет пупочные сосуды от сжатия, тем самым обеспечивая непрерывное поступление питательных веществ и кислорода в эмбрион. Наряду с этим она выполняет защитную функцию, препятствуя проникновению вредоносных агентов из плаценты к эмбриону вне- сосудистым путем.
Для связи зародыша млекопитающих и человека с организмом матери на ранних этапах развития появляются новые провизорные органы – хорион и плацента. Хорион, или ворсинчатая оболочка, развивается из трофобласта и внезародышевой мезодермы. К середине 2-й недели развития зародыш полностью погружен в эндометрий, а трофобласт образует первичные ворсинки – ветвящиеся выпячивания симпласто- и цитоторофобласта, за счет которых после имплантации зародыша в стенку матки устанавливается связь с материнским организмом. С этого момента трофобласт называется уже хорионом (ворсинчатой оболочкой). В начале 3-й недели в ворсины хориона проникает внезародышевая мезенхима (обрастающая изнутри все поверхности, в том числе внутреннюю поверхность трофобласта - хориона). Тем самым формируется соединительнотканная строма ворсин, а сами ворсины становятся вторичными эпителиомезиенхимальными ворсинками хориона.
К концу 3-й недели внутри ворсин из клеток мезенхимы формируются первичные кровеносные сосуды, которые построены как гемокапилляры и вступают в связь с соудами мезодермальной пластинки хориона. Ворсины с развитыми кровеносными сосудами называются третичными и формируют структурно-функциональную единицу плаценты – котиледон. Ворсины хориона погружены в лакуны, образованные в эндометрии и омываются в них материнской кровью, изливающейся из разрушенных сосудов эндометрия. Тем самым устанавливаются маточно-плацентарная циркуляция и обеспечиваемый ею гематотрофный тип питания. Ворсинки хориона разрастаются, ветвятся. При этом на поверхности хориона, обращенной вглубь эндометрия (откуда постоянно поступает материнская кровь), они развиваются и ветвятся более интенсивно, образуя так называемый ветвистый хорион. А на поверхности хориона, обращенной в сторону полости матки, ворсинки постепенно редуцируются, образуя гладкий хорион. Ветвистый хорион вместе с прилежащими к нему участками слизистой оболочки матки образует новый внезародышевый орган - плаценту, обеспечивающую развивающийся зародыш необходимыми питательными веществами и кислородом. Концы ворсин служат своего рода якорями, они прикреплены к эндометрию. В связи с этим, здесь на поверхности ворсин нет симпластотрофобласта, а имеется лишь цитотрофобласт, обеспечивающий плотную фиксацию.
Плаценты разных видов млекопитающих различаются по строению, и в связи с этим, выделяют следующие типы:
1) эпителиохориальная плацента – характеризуется врастанием ворсин хориона в маточные железы и контактом с эпителием этих желез (лошадь, свинья, дельфин, кит), в результате чего обеспечивается поглощение хорионом белков из материнских тканей и расщепление этих белков до аминокислот (синтез собственных белков в эмбрионе происходит в печени);
2) десмохориальная плацента – хорион частично разрушает эпителий маточных желез и ворсины врастают в подлежащую соединительную ткань (парнокопытные млекопитающие - коровы, овцы). Этими типами плацент обеспечивается донашивание зародыша до такого состояния, что к моменту родов он уже способен к самостоятельному питанию и передвижению;
3) эндотелиохориальная плацента – ворсинки хориона разрушают эпителий и соединительную ткань матки и контактируют с эндотелием сосудов (хищные млекопитающие: кошачьи, псовые, тюлени, моржи);
4) гемохориальная плацента – разрушается стенка сосудов матки и ворсинки хориона входят в контакт с материнской кровью (приматы, человек и др.). Эти два типа плацент обеспечивают использование готовых материнских белковдля строительства собственных тканей.
Плацента выполняет трофическую, дыхательную, выделительную, эндокринную, защитную и другие функции. Через плаценту проходят из крови матери аминокислоты, глюкоза, липиды, электролиты, витамины, гормоны, вода, кислород, лекарственные препараты, а также вирусы. Из эмбриона в кровь матери выделяются продукты метаболизма и углекислый газ.
В результате сложных процессов развития к пятой неделе оказываются полностью сформированными все провизорные органы, способные полностью обеспечивать все условия внутриутробного развития организма, который вступает в период органо- и гистогенеза.
ЧАСТНАЯ ГИСТОЛОГИЯ
ЭМБРИОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ И ОРГАНОВ ЧУВСТВ
Нервная ткань развивается из эктодермы. Под индуктивным влиянием нотохорда (растущего вперед) в срединной части эктодермы образуется утолщение, тажке растущее вперед от первичного узелка, – нервная пластинка. На 18-й день начинается прогибание нервной пластинки – появляются нервный желобок и нервные валики. Затем, на протяжении
4-й недели, нервный желобок постепенно замыкается, отделяясь от кожной эктодермы. Однако, смыкание краев желобка и образование нервной трубки происходит не одновременно на всем протяжении: вначале замыкание происходит в шейном отделе, потом – в каудальном и, наконец, в головном отделе зародыша. В результате образуется непарная нервная трубка. При образовании нервного желобка на границе его с кожной эктодермой развиваются утолщения - нервные валики. Группы клеток нервных валиков по мере отделения нервной трубки от кожной эктодермы обособляются и образуют сплошной слой, располагающийся между эктодермой и нервной трубкой – ганглиозную пластинку, или нервные гребни. Часть клеток гиз англиозной пластинки мигрирует в разных направлениях, образуя вегетативные ганглии. Оставшийся материал пластинок сегментируется на спинномозговые узлы.
Таким образом, уже в ранний период развития зародыша обнаруживается разделение нервной системы на три отдела: головной, средний и хвостовой. Наиболее быстро дифференцируется средний отдел, из которого развивается спинной мозг, головной конец служит источником образования головного мозга, который развивается позже, хвостовой конец нервного желобка в нервную трубку превращается позднее выше лежащих отделов и останавливается в своем развитии уже на ранних стадиях, давая хвостовой, слаборазвитый отдел спинного мозга.
РАЗВИТИЕ СПИННОГО МОЗГА
Зачаток спинного мозга имеет вначале вид трубки, которая состоит из слоя клеток цилиндрической формы; последние интенсивно делятся митозом. В результате этого стенка нервной трубки утолщается и приобретает многослойный характер. На поперечном сечении в нервной трубке различают 4 участка стенки: дорсальный - пластинку крыши, вентральный - пластику дна и две боковых пластинки. Утолщенные боковые стенки нервной трубки, а в свою очередь, делятся продольной бороздкой на дорзальную пластинку и вентральную - основную.
Затем наступает дифференцировка клеток нервной трубки в двух направлениях: одни клетки становятся спонгиобластами, образуюшими в дальнейшем нейроглию, другие – нейробластами, превращающимися в дальнейшем в нервные клетки – нейроны.
На этой стадии развития в нервной трубке можно выделить три слоя: 1) внутренний эпендимный слой, выстилающий канал нервной трубки; 2) средний слой - мантийный (плащевой), содержащий нейробласты и дифференцироующиеся из них нейроны, а также примитивные нейроглиальный остов из спонгиобластов (дающих начало клеткам нейроглии – астроцитам и олигодендроцитам); 3) наружный слой - краевую вуаль. Последний слой не содержит нейробластов, образован отростками клеток эпендимного и плащевого слоев, дающим начало проводящим путям мозга, то есть белому веществу. В дальнейшем клетки эпендимного слоя превращаются в цилиндрические эпендимные (глиальные) клетки, выстилающие центральный канал спинного мозга. Из мантийного слоя образуется серое вещество, в котором намечаются группы быстро увеличивающихся в размерах нервных клеток. Это зачатки двигательных ядер. Их нейриты вырастают из спинного мозга на периферию, образуя передние корешки спинномозговых нервов.
В этот период в нервной трубке отмечается неравномерный рост стенок. Пластинка крыши и пластинка дна отстают в росте, сохраняя до конца развития незначительную толщину. Развивающийся спинной мозг, прогибается в области дорзальной и вентральной пластинок, а его правая и левая половины сближаются. В результате чего образуется сзади глиальная и соединительнотканная перегородка, а спереди узкая щель.
В задние рога входят нейриты чувствительных клеток, располагающихся в спинномозговых узлах. Этими нейритами образованы задние корешки спинного мозга.
РАЗВИТИЕ СПИННОМОЗГОВЫХ И ВЕГЕТАТИВНЫХ УЗЛОВ.
Исходным материалом для спинномозговых и вегетативных узлов служит ганглиозная пластинка.
В начале ганглиозная пластинка представлена сплошным клеточным пластом, располагающимся между эктодермой и нервной трубкой. Затем происходит сегментация ганглиозной пластинки вместе с сегментацией зародыша, каждый сегмент ганглиозной пластинки в дальнейшем делится пополам. Эти две части каждого сегмента пластинки и представляют зачатки спинномозговых узлов. В результате интенсивного развития нервной трубки зачатки спинномозговых узлов перемещаются с дорсальной поверхности нервной трубки на ее боковые поверхности, располагаясь по сторонам нервной трубки. Клетки зачатков спинномозговых узлов в процессе эмбриогенеза дифференцируются в двух направлениях: одни клетки превращаются в нейробласты, другие - в спонгиобласты; затем последние развиваются в олигодендроглию ганглия (мантийные клетки). Из нейробластов дифференцируются биполярные нейроны. Биполярные клетки у высших позвоночных животных и человека в процессе созревания изменяют свою форму, отростки клеток постепенно сближаются и их основания сливаются. Таким образом, из тела нейрона идет отросток, который Т - образно делится на 2 отростка - дендрит и нейрит. Так дифференцируются псевдоуниполярные нервные клетки.
Клетки олигодендроглии в дальнейшем образуют капсулу вокруг тел нейронов и оболочки нервных волокон.
Исходным материалом для образования вегетативных нервных узлов служат клеточные элементы ганглиозной пластинки, которые смещаются на периферию в места локализации вегетативных нервных узлов и дифференцируются в двух направлениях: в нейробласты и спонгиобласты. Из нейробластов в последствии образуются мультиполярные нейроны, а из спонгиобластов - клетки глии.
Кроме клеток ганглиев, из ганглиозной пластинки выселяются на периферию также хромаффинобласты, образующие мозговое вещество надпочечников.
ОБРАЗОВАНИЕ ПЕРИФЕРИЧЕСКИХ НЕРВОВ
Периферические отростки псевдоуниполярных клеток, вырастая, сближаются с передними корешками спинного мозга, образуя вместе с ними периферические нервы. Нейриты одного вида мультиполярных нейронов вегетативных узлов входят также в периферические нервы и направляются к органам.
РАЗВИТИЕ ГОЛОВНОГО МОЗГА
В головном отделе зародыша нервная пластинка значительно шире, чем в среднем и в хвостовом отделах. Сворачивание её в желобок и образование нервной трубки происходит медленнее и заканчивается позднее. Вместе с тем, общий рост нервной трубки в головном отделе зародыша совершается неравномерно, вследствие чего в одних участках она оказывается сильно расширенной, а в других - резко суженной. Расширенные участки вначале образуют 3 первичных мозговых пузыря: передний (prosencephalon), средний (mesencephalon) и задний (rhombencephalon). Но вскоре передний мозговой пузырь подразделяется на два: telencephalon и diencephalon. Средний мозговой пузырь остается неразделенным. Задний мозговой пузырь подразделяется на metencephalon и meyelencephalon. Вначале все 5 мозговых пузырей лежат на одной линии, но очень скоро, благодаря интенсивному росту, их взаиморасположение меняется. Появляются 3 изгиба: два изгиба, обращенных кзади - теменной (на уровне среднего мозга) и затылочный (на границе между задним мозговым пузырем и спинным мозгом), и один, обращенный кпереди - мостовой (на уровне передней части заднего мозгового пузыря).
Дальнейшее развитие отделов мозга идет различно потому, что в разных мозговых пузырях рост их стенки совершается неодинаково. В связи с этим, в стенах мозговых пузырей образуются в одних случаях глубокие складки, с появлением которых связан процесс возникновения в глубине некоторых отделов мозга ядер серого вещества. В других случаях образуются более мелкие поверхностные складки, обуславливающие появление на поверхности некоторых мозговых пузырей множественных борозд и извилин.
Особенно интенсивно развивается передний мозговой пузырь. Вначале он представляет собой непарное образование, затем от передних отделов боковых его стенок образуется с каждой стороны по небольшому выпячиванию являющимися зачатками больших полушарий мозга. В то же время спереди сверху из окружающей мезенхимы по средней линии между ними врастает соединительнотканная перегородка, разделяющая пузырь на 2 половины. Полость пузыря при этом преобразуется в два боковых желудочка.
В дальнейшем, закладки больших полушарий достигают максимального объёма (5-6 месяц), превышающего объём производных других четырёх мозговых пузырей. На поверхности полушарий образуются борозды и извилины, так же происходит разделение полушарий на отдельные доли. На медиальных поверхностях развиваются утолщения, которые срастаются между собой (вследствие прорастания нервных волокон) и образуют мозолистое тело, связывающее эти полушария друг с другом. Следует отметить, что уже в первые месяцы развития на передних поверхностях больших полушарий, в виде растущих кпереди выпячиваний, образуется первая пара черепно-мозговых нервов - обонятельные нервы, которые вступают в связь с чувствительны эпителием слизистой оболочки обонятельного отдела носовой полости.
Второй мозговой пузырь вначале является самым крупным, в последующем растет медленно. На ранних стадиях развития на его боковых стенках в виде выпячиваний образуются глазные пузыри, ножки которых дают вторую пару черепно-мозговых нервов - зрительные нервы. У основания глазных пузырей боковые стенки разрастаются, превращаясь в зрительные бугры. Также, путем выпячивания, из дорзальной стенки второго мозгового пузыря образуется эпифиз (шишковидная железа), а из вентральной (в виде воронки) - задняя доля гипофиза (нейрогипофиз). Из утолщения задней стенки этого пузыря развивается серый бугор и сосцевидные тела. Полость второго мозгового пузыря сохраняется в виде третьего желудочка.
Третий мозговой пузырь развивается незначительно. За счет утолщающихся пластинок дна и нижних отделов боковых пластинок образуются ножки мозга. Верхние части боковых пластинок при своем развитии превращаются в четверохолмие. За счет пластинки крыши развивается передний мозговой парус. Полость третьего мозгового пузыря, вследствие разрастания всех его стенок, сильно суживается, сохраняясь в виде протока, так называемого сильвиевого водопровода.
Четвертый мозговой пузырь развивается таким образом, что его боковые пластики значительно разрастаются, а пластинки крыши и дна редуцируются. При этом на верхней части боковых пластинок образуется мозжечок, а из нижних частей их - варолиев мост. Полость пузыря резко суживается и представляет собой в последующем передний отдел четвертого желудочка, тогда как основная часть его образуется из полости пятого мозгового пузыря.
Пятый мозговой пузырь - идет на построение продолговатого мозга. При этом разрастаются лишь бо

