




Основными показателями структуры популяций является численность и распределение организмов в пространстве и соотношение разнокачественных особей. В связи с размерами ареала популяций может значительно изменяться и численность особей в популяциях.
Численность популяции — это общее количество особей на данной территории или в данном объеме. Зависит от соотношения интенсивности размножения (плодовитости) и смертности. В период размножения происходит рост популяции. Смертность же, наоборот, приводит к сокращению ее численности.
Плотность популяции определяется количеством особей или биомассой на единицу площади либо объема, например: 400 деревьев на 1 га, 0,5 г циклопов в 1 м3 воды. Нередко важно различать среднюю плотность, т. е. численность или биомассу на единицу всего пространства, кудельную или экологическую плотность — численность или биомассу на единицу обитаемого пространства, доступной площади объема, которые фактически могут быть заняты популяцией.
Плотность популяции отличается изменчивостью и зависит от ее численности. При возрастании численности не наблюдается увеличение плотности лишь в том случае, когда возможно распределение популяции, расширение ее ареала. Особи, составляющие популяции, имеют различные типы пространственного распределения, выражающие их реакции на различные влияния, например, добычу и благоприятные физические условия или конкурентные реакции. Различают три типа распределения или расселения особей внутри популяции: равномерное, случайное и групповое (рис. 9.2).
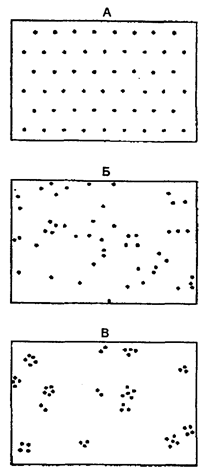
Рис. 9.2. Основные типы распределения особей в популяции:
А — равномерное распределение; Б – случайное распределение; В - групповое распределение (по Одуму, 1986)
Равномерное распределение в природе чаще связано с острой конкуренцией между разными особями. Такой тип распределения отмечают у хищных рыб и у колюшек с их территориальным инстинктом и сугубо индивидуальным характером.
Случайное распределение имеет место только в однородной среде. Так на первых порах распределяется тля на поле. По мере ее размножения распределение приобретает групповой или пятнистый (конгрегационный) характер.
Групповое распределение встречается наиболее часто. Так, в сосновом лесу деревья вначале расселяются группами, а в дальнейшем их размещение становится равномерным. Популяции групповое распределение обеспечивает (по Ю. Одуму, 1986) более высокую устойчивость по отношению к неблагоприятным условиям по сравнению с отдельной особью. Животные, ведущие подвижный образ жизни, как правило, распределяются активно, что приводит к интенсивному перемешиванию популяций и стиранию границ между ними. Например, очень подвижные и активно перемещающиеся песцы, другие животные, птицы имеют огромные ареалы без резких границ между популяциями.
У пассивно передвигающихся и малоподвижных организмов, наоборот, популяции четко разграничены даже на относительно небольшой территории. Таковы популяции наземных моллюсков, многих земноводных. Размеры ареала популяции зависят от величины особей, составляющей ее. Мелкие особи занимают сравнительно небольшие ареалы, тогда как у видов с крупными особями они обширны. Вместе с тем это правило имеет много исключений. Так, территория, занимаемая популяцией прыткой ящерицы Lacerta agilis, может колебаться от 0,1 до нескольких гектаров.
Знание типа распределения организмов имеет большое значение при оценке плотности популяции методом выборки (в случае группового размещения площадь выборки должна быть большая). Возьмем n выборок. Среднее число особей в каждой выборке обозначим через m и получим рассеяние или дисперсию S2 по формуле:
 (9.1)
(9.1)
При равномерном распределении дисперсия S2 равна нулю, так как число особей в каждой выборке постоянно и равно среднему. В случае случайного распределения среднее m и дисперсия S2 равны. При групповом распределении рассеяние S2 выше среднего и разница между ними тем больше, чем сильнее тенденция животных к образованию скоплений.
Рождаемость и смертность
Динамика численности и плотности популяций находится в тесной зависимости от рождаемости или плодовитости и смертности.
Рождаемость — это способность популяции к увеличению численности. Характеризует частоту появления новых особей в популяции. Различают рождаемость абсолютную и удельную. Абсолютная (общая) рождаемость — число новых особей (DNn), появившихся за единицу времени (Dt). Удельная рождаемость выражается в числе особей на особь в единицу времени:
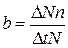 . (9.2)
. (9.2)
Так, для популяций человека как показатель удельной рождаемости используют число детей, родившихся в год на 1000 человек. В живых организмах заложена огромная возможность к размножению и подтверждается правилом максимальной рождаемости (воспроизводства): в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей. Оно достигается в идеальных условиях, когда отсутствуют лимитирующие экологические факторы и размножение ограничено лишь физиологическими особенностями вида. Например, один одуванчик менее чем за 10 лет способен заселить своими потомками земной шар, если все семена прорастут. Другой пример. Бактерии делятся каждые 20 мин. При таком темпе одна клетка за 36 ч может дать потомство, которое покроет сплошным слоем всю нашу планету. Обычно же существует экологическая или реализуемая рождаемость, возникающая в обычных или специфических условиях среды. Средняя величина плодовитости выработана исторически как приспособление, которое обеспечивает пополнение убыли популяций. Естественно, что у менее приспособленных видов к неблагоприятным условиям высокая смертность в молодом (личиночном) возрасте компенсируется значительной плодовитостью.
Среди насекомых самая высокая плодовитость у растительноядных форм, а низкая — у хищников и паразитов. В благоприятных условиях плодовитость, как правило, низкая. Характер плодовитости зависит и от скорости полового созревания, числа генераций в течение сезона, от состояния в популяции самок и самцов. Если вид размножается с большой скоростью и чутко реагирует на изменения условий среды, то численность популяций его быстро и существенно изменяется. Это относится к многим насекомым и мышевидным грызунам. Таким образом, максимальная рождаемость или плодовитость является константой, определяемой расчетным путем, например, умножением среднего числа гнезд, которое способна построить самка птицы за год, на такое же число яиц, которые она может отложить в наиболее благоприятную часть сезона года. Максимальная рождаемость — тот предел, который характерен для скоростей увеличения числа особей в популяции. Правило максимальной рождаемости (воспроизводства) есть частный случай закона максимума биогенной энергии (энтропии) В. И. Вернадского — Э. С. Бауэра.
Численность и плотность популяции зависит и от ее смертности. Смертность популяции — это количество особей, погибших за определенный период. Абсолютная (общая) смертность — это число особей, погибших в единицу времени (DNm).
Удельная смертность (d) выражается отношением абсолютной смертности к численности популяции:

Абсолютная и удельная смертность характеризуют скорость убывания численности популяции вследствие гибели особей от хищников, болезней, старости и т. д.
Различают три типа смертности. Первый тип смертности характеризуется одинаковой смертностью во всех возрастах. Выражается экспоненциальной кривой (убывающей геометрической прогрессии). Данный тип смертности встречается редко и только у популяций, которые постоянно находятся в оптимальных условиях.
Второй тип смертности характеризуется повышенной гибелью особей на ранних стадиях развития и свойствен большинству растений и животных. Максимальная гибель животных происходит в личиночной фазе или в молодом возрасте, у многих растений — в стадии произрастания семян и всходов. У насекомых до взрослых особей доживает 0,3—0,5% отложенных яиц, у многих рыб — 1—2% количества выметанной икры.
Третий тип смертности отличается повышенной гибелью взрослых, в первую очередь старых, особей. Отличается он у насекомых, личинки которых обитают в почве, воде, древесине, а также в других местах с благоприятными условиями. В экологии широкое распространение получило графическое построение «кривых выживания» (рис. 9.3).

Рис. 9.3. Различные типы кривых выживания
Откладывая по оси абсцисс продолжительность жизни в процентах от общей продолжительности жизни, можно сравнивать кривые выживания организмов, продолжительность жизни которых имеет значительные различия. На основании таких кривых можно определить периоды, в течение которых тот или иной вид особенно уязвим. Поскольку смертность подвержена более резким колебаниям и больше зависит от факторов окружающей среды, чем рождаемость, она играет главную роль в регулировании численности популяции.

