




В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
17. Хроматин (греч. chroma — цвет, краска и греч. nitos — нить) — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, — надмолекулярных структур, участвующих в упаковке хромосом.
1) эухроматин, локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компакт-ный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка.
2) Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. гетерохроматин - плотно спирализованная часть хроматина. Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окра-шивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул.
18. Митотический и жизненный циклы. Период существования клетки от момента ее образования путем деления материнской клетки (включая само деление) до собственного деления или смерти называют жизненным (клеточным) циклом.
Продолжительность жизненного цикла различных клеток многоклеточного организма различна. Так, клетки нервной ткани после завершения эмбрионального периода перестают делиться и функционируют на протяжении всей жизни организма, а затем погибают. Клетки же зародыша на стадии дробления, завершив одно деление, сразу приступают к следующему, минуя все остальные фазы.
Митоз – непрямое деление соматических клеток, в результате которого сначала происходит удвоение, а затем равномерное распределение наследственного материала между дочерними клетками.
Биологическое значение митоза: в результате митоза образуются две клетки, каждая из которых содержит столько же хромосом, сколько их было в материнской. Дочерние клетки генетически идентичны родительской. Число клеток в организме увеличивается, что представляет собой один из главных механизмов роста. Многие виды растений и животных размножаются бесполым путем при помощи одного лишь митотического деления клеток, таким образом, митоз лежит в основе размножения. Митоз обеспечивает регенерацию утраченных частей и замещение клеток, происходящее в той или иной степени у всех многоклеточных организмов.
Митотический цикл состоит из интерфазы и митоза. Длительность митотического цикла у разных организмов сильно варьирует. Непосредственно на деление клетки уходит обычно 1–3 ч, то есть основную часть жизни клетка находится в интерфазе.
Интерфазой называют промежуток между двумя клеточными делениями. Продолжительность интерфазы, как правило, составляет до 90% всего клеточного цикла. Интерфаза состоит из трех периодов: пресинтетический, или G1; синтетический, или S; постсинтетический, или G2.
Начальный отрезок интерфазы – пресинтетический период (2n2с, где n – количество хромосом, с – количество ДНК), период роста, начинающийся непосредственно после митоза. Синтетический период по продолжительности очень различен: от нескольких минут у бактерий до 6–12 ч в клетках млекопитающих. Во время синтетического периода происходит самое главное событие интерфазы – удвоение молекул ДНК. Каждая хромосома становится двухроматидной, а число хромосом не изменяется (2n4с).
Постсинтетический период. Обеспечивает подготовку клетки к делению и также характеризуется интенсивными процессами синтеза белков, входящих в состав хромосом; синтезируются ферменты и энергетические вещества, необходимые для обеспечения процесса деления клетки.
Митоз. Для удобства изучения происходящих во время деления событий митоз разделяют на четыре стадии: профазу, метафазу, анафазу, телофазу.
Профаза (2n4с). В результате спирализации хромосомы уплотняются, укорачиваются. В поздней профазе хорошо видно, что каждая хромосома состоит из двух хроматид, соединенных центромерой. Хромосомы начинают передвигаться к клеточному экватору. Формируется веретено деления, ядерная оболочка исчезает, и хромосомы свободно располагаются в цитоплазме. Ядрышко обычно исчезает чуть раньше.
Метафаза (2n4с). Хромосомы выстраиваются в плоскости экватора, образуя так называемую метафазную пластинку. Центромеры хромосом лежат строго в плоскости экватора. Нити веретена прикрепляются к центромерам хромосом, некоторые нити проходят от полюса к полюсу клетки, не прикрепляясь к хромосомам.
Анафаза (4n4с). Начинается с деления центромер всех хромосом, в результате чего хроматиды превращаются в две совершенно обособленные, самостоятельные дочерние хромосомы. Затем дочерние хромосомы начинают расходиться к полюсам клетки.
Репарация ДНК
Процесс, позволяющий живым организмам восстанавливать повреждения, возникающие в ДНК, называют репарацией. Все репарационные механизмы основаны на том, что ДНК - двухцепочечная молекула, т.е. в клетке есть 2 копии генетической информации. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена.Процесс репарации происходит в несколько этапов. На первом этапе выявляется нарушение комплементарности цепей ДНК. В ходе второго этапа некомплементарный нуклеотид или только основание устраняется, на третьем и четвёртом этапах идёт восстановление целостности цепи по принципу комплементарности. Однако в зависимости от типа повреждения количество этапов и ферментов, участвующих в его устранении, может быть разным.Очень редко происходят повреждения, затрагивающие обе цепи ДНК, т.е. нарушения структуры нуклеотидов комплементарной пары. Такие повреждения в половых клетках не репарируются, так как для осуществления сложной репарации с участием гомологичной рекомбинации требуется наличие диплоидного набора хромосом.
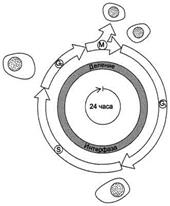
Рис. 4-20. Фазы клеточного цикла. После фазы М, в ходе которой происходит деление ядра (митоз) и цитоплазмы (цитокинез), дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы G1, в ходе которой активно происходят биосинтетические процессы, резко замедленные во время митоза. Фаза S - период синтеза ДНК; она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются. Затем наступает фаза G2, в ходе которой происходят деление митохондрий и увеличение энергетических запасов клетки. Фаза G2продолжается до начала митоза, т.е. фазы М. В фазе М ядерная оболочка разрушается, формируются два новых ядра, цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. На рисунке представлен 24-часовой цикл.
А. Спонтанные повреждения
Нарушения комплементарности цепей ДНК могут происходить спонтанно, т.е. без участия каких-либо повреждающих факторов, например
Таблица 4-2. Циклимы и циклинзависимые киназы, регулирующие прохождение клеточного цикла
| Циклин | Киназа | Функция |
| D,E | CDK4, CDK6 | Регулирует переход клетки из G1-фазы в S-фазу |
| А | CDK2 | Активирует синтез ДНК на начальной стадии S-фазы |
| В | CDK1 | Регулирует переход клетки из G2-фазы в М-фазу |
в результате ошибок репликации, дезаминирования нуклеотидов, депуринизации.
Ошибки репликации
Точность репликации ДНК очень велика, но примерно один раз на 105-106 нуклеотидных остатков происходят ошибки спаривания, и тогда вместо пары нуклеотидов А-Т, G-С в дочернюю цепь ДНК оказываются включёнными нук-леотиды, некомплементарные нуклеотидам матричной цепи. Однако ДНК-полимеразы д, е способны после присоединения очередного нук-леотида в растущую цепь ДНК делать шаг назад (в направлении от 3'- к 5'- концу) и вырезать последний нуклеотид, если он некомплементарен нуклеотиду в матричной цепи ДНК. Этот процесс исправления ошибок спаривания (или коррекция) иногда не срабатывает, и тогда в ДНК по окончании репликации остаются некомплементарные пары, тем более, что ДНК-полимераза а лишена корректирующего механизма и "ошибается" чаще, чем другие полимеразы.
При неправильном спаривании в первичной структуре дочерней цепи ДНК необычные основания не появляются, нарушена только ком-плементарность. Система репарации некомплементарных пар должна происходить только на дочерней цепи и производить замену некомплементарных оснований только в ней. Ферменты, участвующие в удалении неправильной пары нуклеотидов, распознают матричную цепь по наличию метилированных остатков аденина в последовательностях -GATC-. Пока основания нуклеотидных остатков в дочерней цепи неметилированы, ферменты должны успеть выявить ошибку репликации и устранить её.
Распознавание и удаление (первый этап) некомплементарного нуклеотида происходят при участии специальных белков mut S, mut L, mut H. Каждый из белков выполняет свою специфическую функцию. Mut S находит неправильную пару и связывается с этим фрагментом. Mut Н присоединяется к метилированному (по аденину) участку -GATC-, расположенному вблизи некомплементарной пары. Связующим между mut S и mut Н служит белок mut L, его присоединение завершает образование активного фермента. Формирование комплекса mut S, mut L, mut Н на участке, содержащем ошибку, способствует проявлению у белка mut Н эндонуклеазной активности. Ферментативный комплекс гидролизует фосфоэфирную связь в неметилированной цепи (рис. 4-21).
К свободным концам цепи присоединяется экзонуклеаза (второй этап). Отщепляя по одному нуклеотиду в направлении от 3'- к 5'- концу дочерней цепи, она устраняет участок, содержащий некомплементарную пару. Брешь застраивает ДНК-полимераза в (третий этап), соединение основного и вновь синтезированного участков цепи катализирует фермент ДНК-лигаза (четвёртый этап). Для успешного функционирования экзонуклеазы, ДНК-полимеразы р и ДНК-лигазы необходимо участие в репарации хеликазы и SSB-белков.

